

Reduction of endotoxicity in Bordetella bronchiseptica by lipid A engineering: Characterization of lpxL1 and pagP mutants
- PMID: 34053396
- PMCID: PMC8168481
- DOI: 10.1080/21505594.2021.1929037
Reduction of endotoxicity in Bordetella bronchiseptica by lipid A engineering: Characterization of lpxL1 and pagP mutants
Abstract
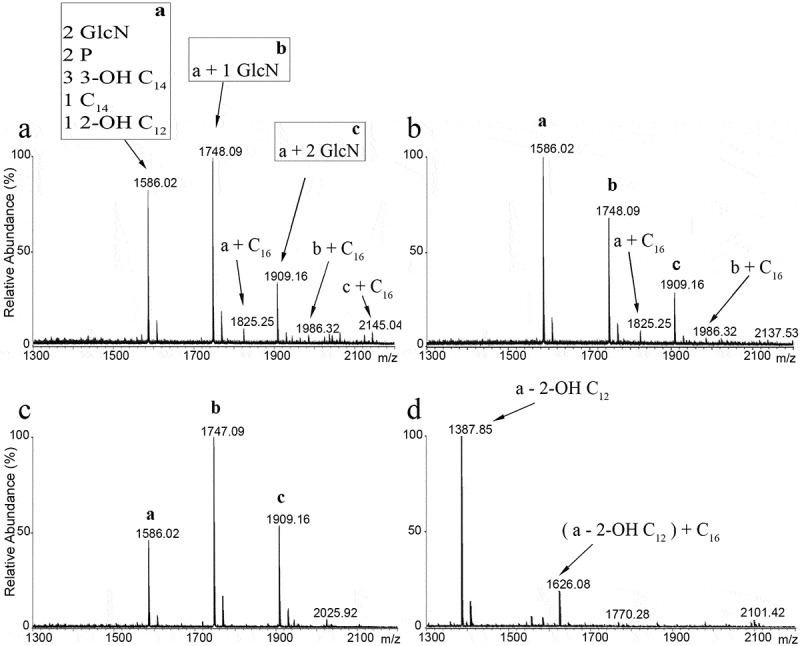
Whole-cell vaccines against Gram-negative bacteria commonly display high reactogenicity caused by the endotoxic activity of lipopolysaccharide (LPS), one of the major components of the bacterial outer membrane. Underacylation of the lipid A moiety of LPS has been related with reduced endotoxicity in several Gram-negative species. Here, we evaluated whether the inactivation of two genes encoding lipid A acylases of Bordetella bronchiseptica, i.e. pagP and lpxL1, could be used for the development of less reactogenic vaccines against this pathogen for livestock and companion animals. Inactivation of pagP resulted in the loss of the secondary palmitate chain at position 3' of lipid A, but hardly affected the potency of the LPS to activate the Toll-like receptor 4 (TLR4). Inactivation of lpxL1 resulted in the loss of the secondary 2-hydroxy laurate group present at position 2 of lipid A and, unexpectedly, in the additional loss of the glucosamines that decorate the phosphate groups at positions 1 and 4' and in an increase in LPS molecules carrying O-antigen. The resulting LPS showed greatly reduced potency to activate TLR4 in HEK-Blue reporter cells expressing human or mouse TLR4 as well as in porcine macrophages. Characterization of the lpxL1 mutant revealed many pleiotropic phenotypes, including increased resistance to SDS and rifampicin, increased susceptibility to cationic antimicrobial peptides, decreased auto-aggregation and biofilm formation, and a tendency to decreased infectivity of macrophages, which are all related to the altered LPS structure. We suggest that the lpxL1 mutant will be useful for the generation of safer vaccines.
Keywords: Bordetella; LPS; LpxL1; PagP; TLR4; antimicrobial susceptibility; autoaggregation; biofilms; endotoxicity; vaccine.
Figures








References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous