

Peptidomics of Haemonchus contortus
- PMID: 34056183
- PMCID: PMC8153747
- DOI: 10.1021/acsomega.1c00650
Peptidomics of Haemonchus contortus
Abstract
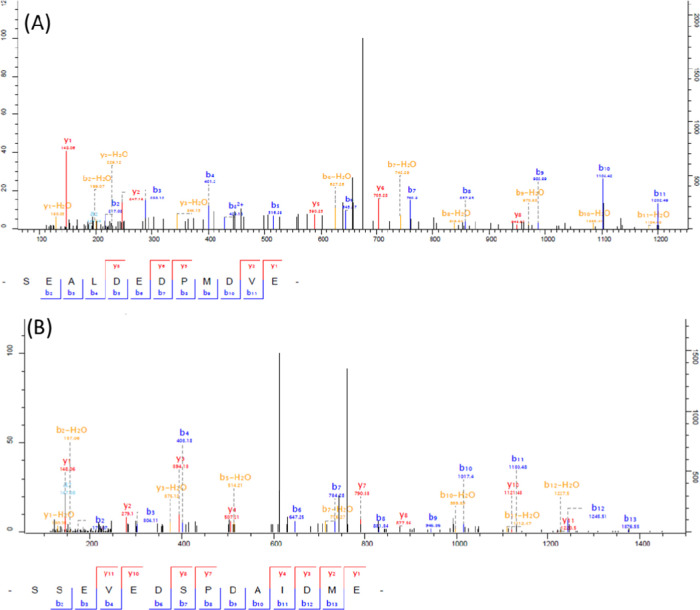
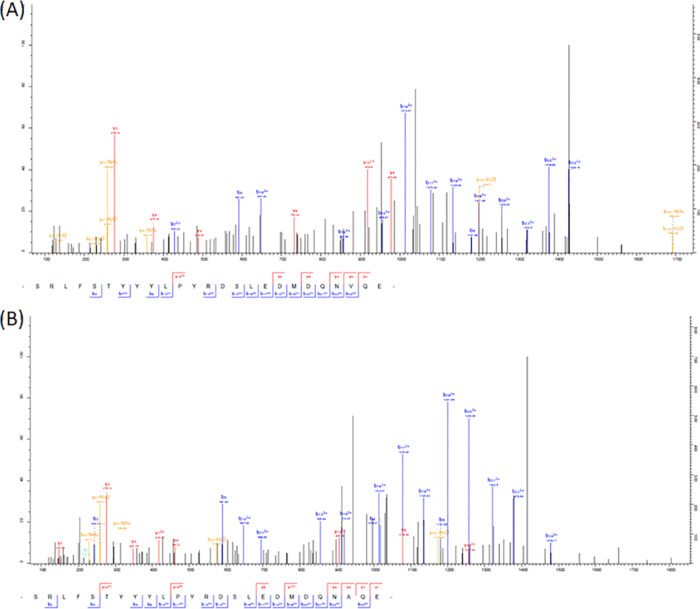
The nematode Haemonchus contortus (the barber's pole worm) is an endoparasite infecting wild and domesticated ruminants worldwide. Widespread anthelmintic resistance of H. contortus requires alternative strategies to control this parasite. Neuropeptide signaling represents a promising target for anthelmintic drugs. Identification and relative quantification of nematode neuropeptides are, therefore, required for the development of such therapeutic targets. In this work, we undertook the profiling of the whole H. contortus larvae at different stages for the direct sequencing of the neuropeptides expressed at low levels in these tissues. We set out a peptide extraction protocol and a peptidomic workflow to biochemically characterize bioactive peptides from both first-stage (L1) and third-stage larvae (L3) of H. contortus. This work led to the identification and quantification at the peptidomic level of more than 180 mature neuropeptides, including amidated and nonamidated peptides, arising from 55 precursors of H. contortus. The differential peptidomic approach provided evidence that both life stages express most FMRFamide-like peptides (FLPs) and neuropeptide-like proteins (NLPs). The H. contortus peptidome resource, established in this work, could add the discovery of neuropeptide system-targeting drugs for ruminants.
© 2021 The Authors. Published by American Chemical Society.
Conflict of interest statement
The authors declare no competing financial interest.
Figures






References
-
- Besier R. B.; Kahn L. P.; Sargison N. D.; Van Wyk J. A. The Pathophysiology, Ecology and Epidemiology of Haemonchus contortus Infection in Small Ruminants. Adv. Parasitol. 2016, 93, 95–143. - PubMed
-
- Gasser R. B.; von Samson-Himmelstjerna G. Haemonchus contortus and haemonchosis– past, present and future trends. Adv. Parasitol. 2016, 93, 1–666.
-
- Veglia E. The anatomy and life history of the Haemonchus contortus. Rep. Dir. Vet. Res. 1915, 3–4, 347–500.
-
- Brown L. A.; Jones A. K.; Buckingham S. D.; Mee C. J.; Sattelle D. B. Contributions from Caenorhabditis elegans functional genetics to antiparasitic drug target identification and validation: nicotinic acetylcholine receptors, a case study. Int. J. Parasitol. 2006, 36, 617–624. 10.1016/j.ijpara.2006.01.016. - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
