

The structure of the mouse ADAT2/ADAT3 complex reveals the molecular basis for mammalian tRNA wobble adenosine-to-inosine deamination
- PMID: 34057470
- PMCID: PMC8216470
- DOI: 10.1093/nar/gkab436
The structure of the mouse ADAT2/ADAT3 complex reveals the molecular basis for mammalian tRNA wobble adenosine-to-inosine deamination
Abstract
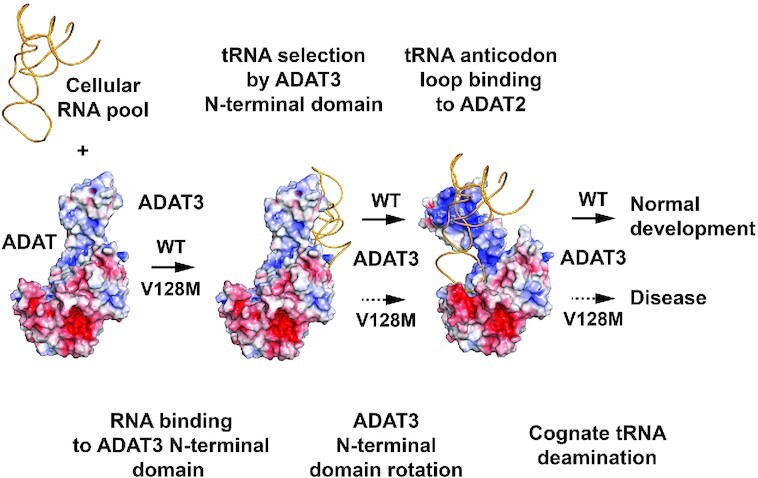
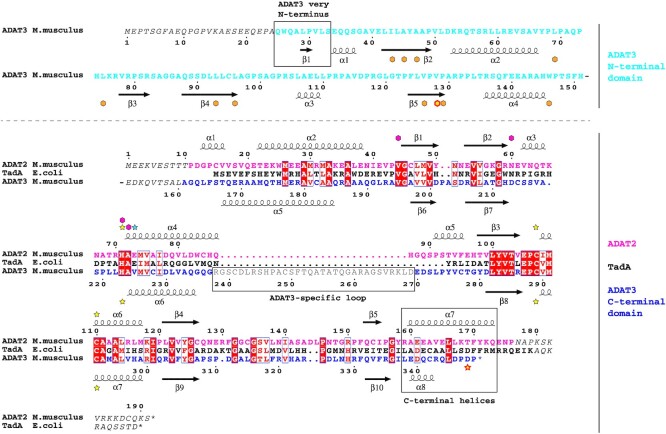
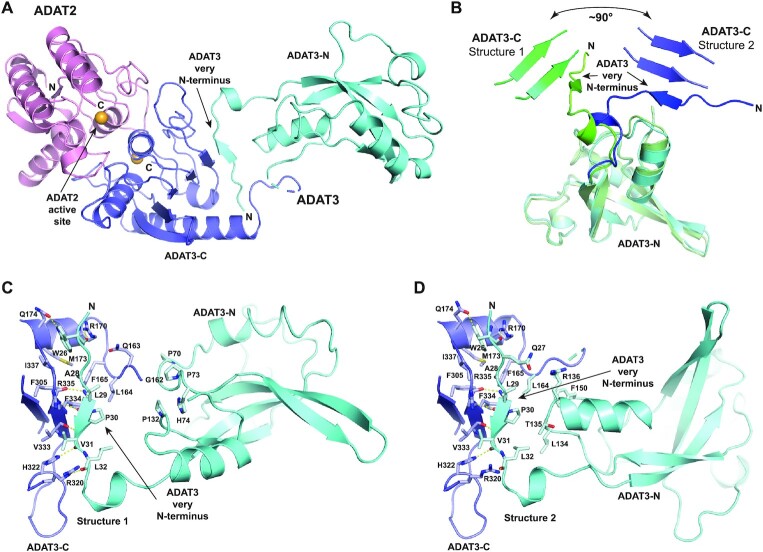
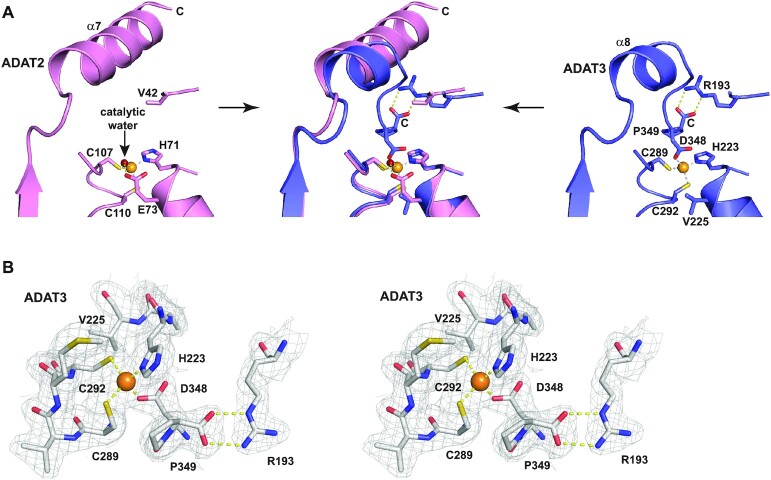
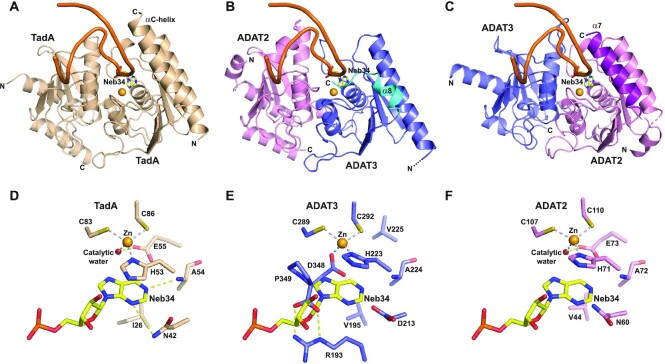
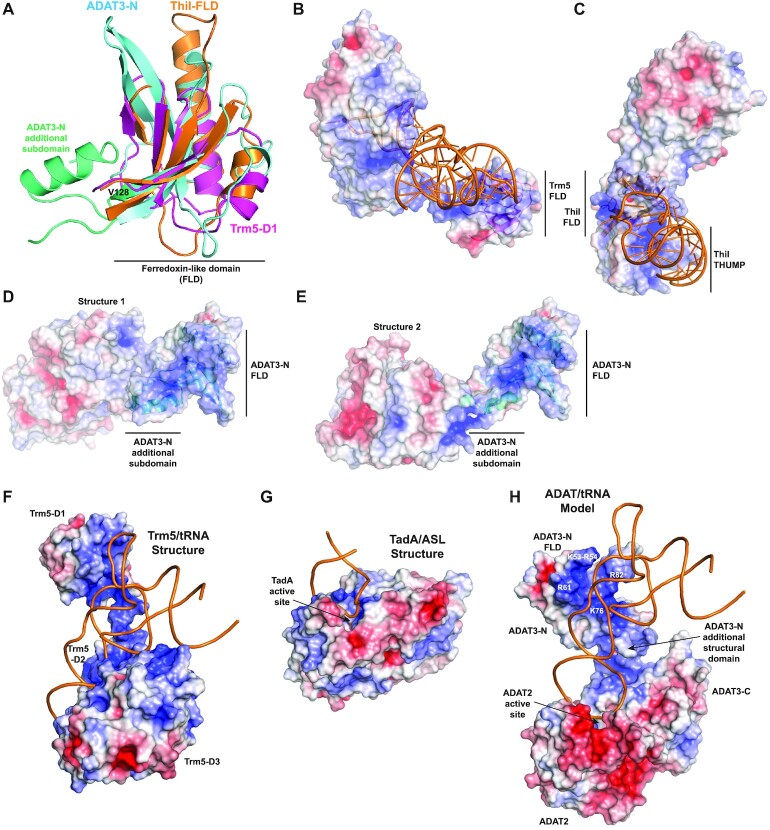
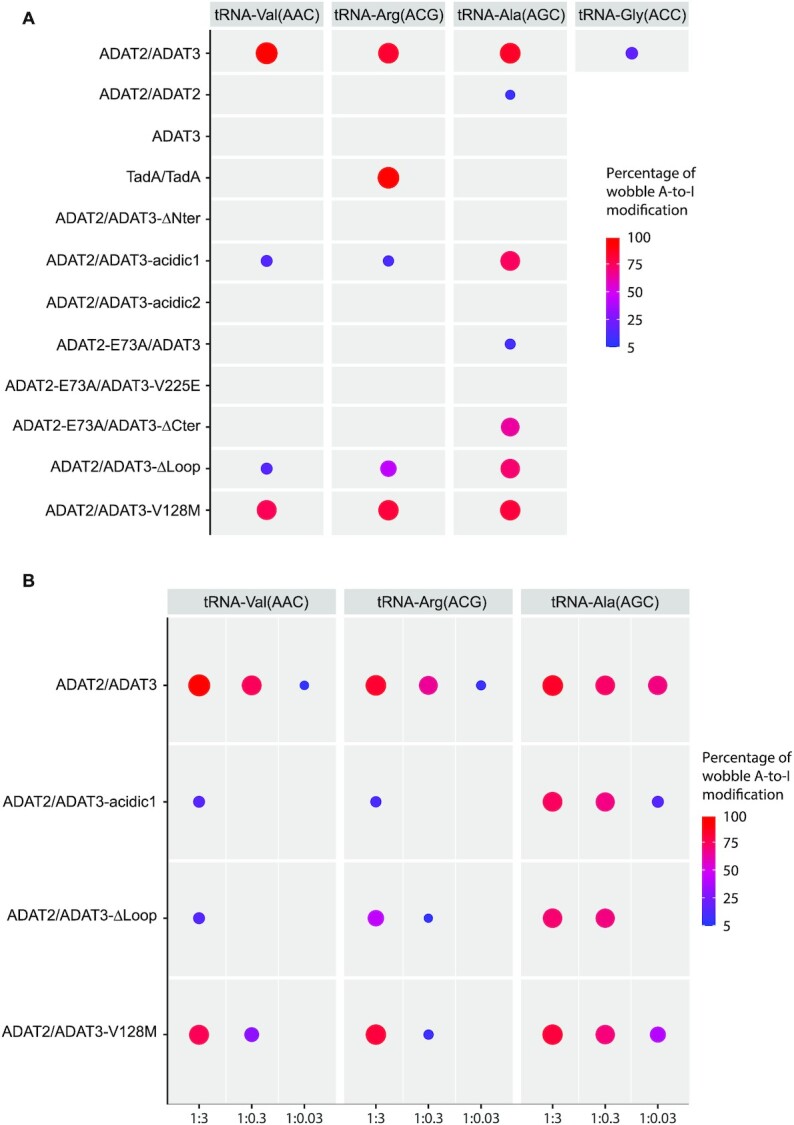
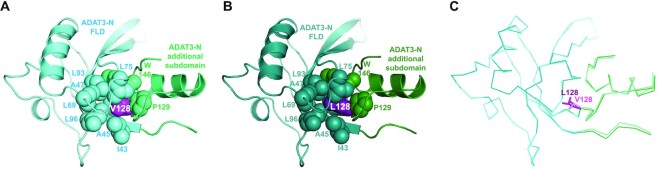
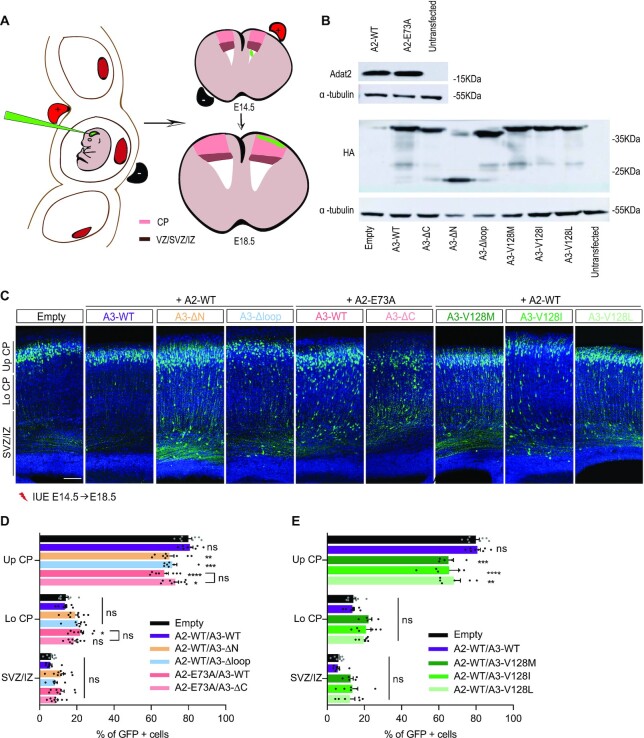
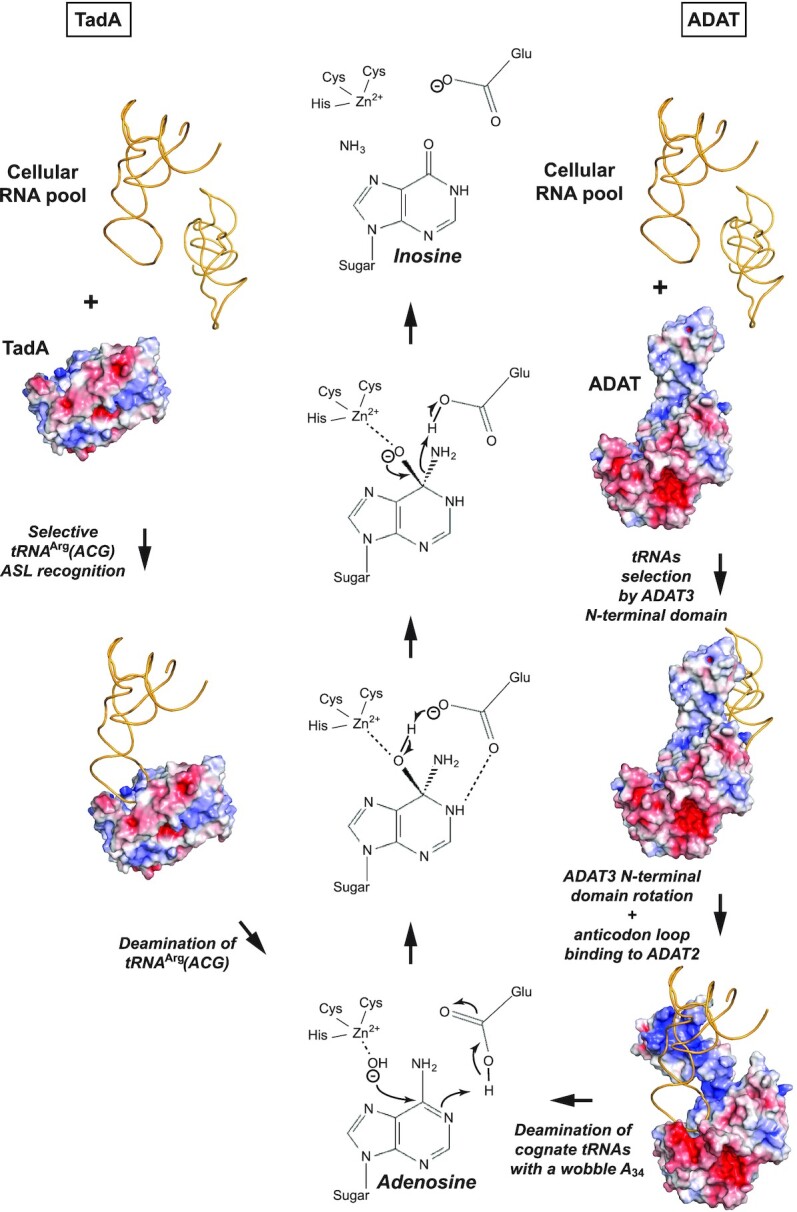
Post-transcriptional modification of tRNA wobble adenosine into inosine is crucial for decoding multiple mRNA codons by a single tRNA. The eukaryotic wobble adenosine-to-inosine modification is catalysed by the ADAT (ADAT2/ADAT3) complex that modifies up to eight tRNAs, requiring a full tRNA for activity. Yet, ADAT catalytic mechanism and its implication in neurodevelopmental disorders remain poorly understood. Here, we have characterized mouse ADAT and provide the molecular basis for tRNAs deamination by ADAT2 as well as ADAT3 inactivation by loss of catalytic and tRNA-binding determinants. We show that tRNA binding and deamination can vary depending on the cognate tRNA but absolutely rely on the eukaryote-specific ADAT3 N-terminal domain. This domain can rotate with respect to the ADAT catalytic domain to present and position the tRNA anticodon-stem-loop correctly in ADAT2 active site. A founder mutation in the ADAT3 N-terminal domain, which causes intellectual disability, does not affect tRNA binding despite the structural changes it induces but most likely hinders optimal presentation of the tRNA anticodon-stem-loop to ADAT2.
© The Author(s) 2021. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Similar articles
-
ADATs: roles in tRNA editing and relevance to disease.Acta Biochim Biophys Sin (Shanghai). 2024 Jul 22;57(1):73-83. doi: 10.3724/abbs.2024125. Acta Biochim Biophys Sin (Shanghai). 2024. PMID: 39034823 Free PMC article. Review.
-
Identification and rescue of a tRNA wobble inosine deficiency causing intellectual disability disorder.RNA. 2020 Nov;26(11):1654-1666. doi: 10.1261/rna.076380.120. Epub 2020 Aug 6. RNA. 2020. PMID: 32763916 Free PMC article.
-
ADAT3 variants disrupt the activity of the ADAT tRNA deaminase complex and impair neuronal migration.Brain. 2025 Sep 3;148(9):3407-3421. doi: 10.1093/brain/awaf109. Brain. 2025. PMID: 40120092 Free PMC article.
-
Formation of tRNA Wobble Inosine in Humans Is Disrupted by a Millennia-Old Mutation Causing Intellectual Disability.Mol Cell Biol. 2019 Sep 11;39(19):e00203-19. doi: 10.1128/MCB.00203-19. Print 2019 Oct 1. Mol Cell Biol. 2019. PMID: 31263000 Free PMC article.
-
Enzymatic conversion of adenosine to inosine and to N1-methylinosine in transfer RNAs: a review.Biochimie. 1996;78(6):488-501. doi: 10.1016/0300-9084(96)84755-9. Biochimie. 1996. PMID: 8915538 Review.
Cited by
-
ADATs: roles in tRNA editing and relevance to disease.Acta Biochim Biophys Sin (Shanghai). 2024 Jul 22;57(1):73-83. doi: 10.3724/abbs.2024125. Acta Biochim Biophys Sin (Shanghai). 2024. PMID: 39034823 Free PMC article. Review.
-
Engineered deaminases as a key component of DNA and RNA editing tools.Mol Ther Nucleic Acids. 2023 Oct 20;34:102062. doi: 10.1016/j.omtn.2023.102062. eCollection 2023 Dec 12. Mol Ther Nucleic Acids. 2023. PMID: 38028200 Free PMC article. Review.
-
Amphioxus adenosine-to-inosine tRNA-editing enzyme that can perform C-to-U and A-to-I deamination of DNA.Commun Biol. 2023 Jul 18;6(1):744. doi: 10.1038/s42003-023-05134-0. Commun Biol. 2023. PMID: 37464027 Free PMC article.
-
Antibody production relies on the tRNA inosine wobble modification to meet biased codon demand.Science. 2024 Jan 12;383(6679):205-211. doi: 10.1126/science.adi1763. Epub 2024 Jan 11. Science. 2024. PMID: 38207021 Free PMC article.
-
RNA editing enzymes: structure, biological functions and applications.Cell Biosci. 2024 Mar 16;14(1):34. doi: 10.1186/s13578-024-01216-6. Cell Biosci. 2024. PMID: 38493171 Free PMC article. Review.
References
-
- El Yacoubi B., Bailly M., de Crecy-Lagard V.. Biosynthesis and function of posttranscriptional modifications of transfer RNAs. Annu. Rev. Genet. 2012; 46:69–95. - PubMed
-
- Kirchner S., Ignatova Z.. Emerging roles of tRNA in adaptive translation, signalling dynamics and disease. Nat. Rev. Genet. 2015; 16:98–112. - PubMed
-
- Schimmel P. The emerging complexity of the tRNA world: mammalian tRNAs beyond protein synthesis. Nat. Rev. Mol. Cell Biol. 2018; 19:45–58. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
