

Mutation of SlARC6 leads to tissue-specific defects in chloroplast development in tomato
- PMID: 34059665
- PMCID: PMC8167136
- DOI: 10.1038/s41438-021-00567-2
Mutation of SlARC6 leads to tissue-specific defects in chloroplast development in tomato
Abstract
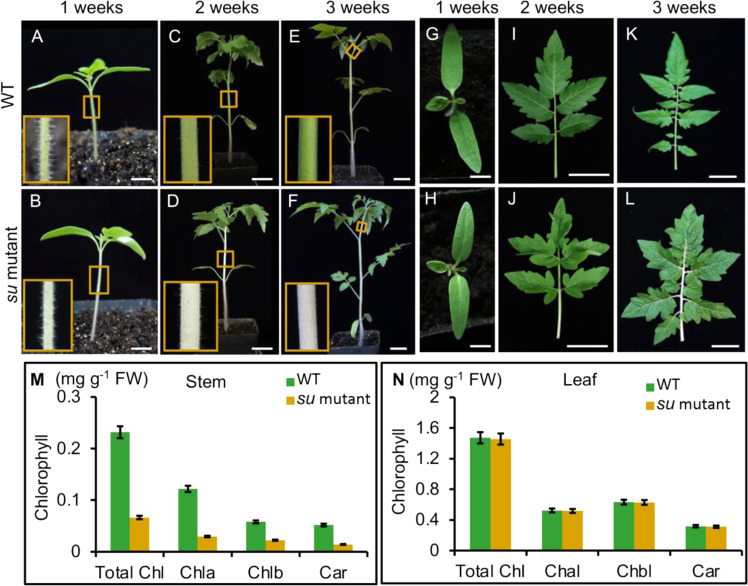
The proliferation and development of chloroplasts are important for maintaining the normal chloroplast population in plant tissues. Most studies have focused on chloroplast maintenance in leaves. In this study, we identified a spontaneous mutation in a tomato mutant named suffulta (su), in which the stems appeared albinic while the leaves remained normal. Map-based cloning showed that Su encodes a DnaJ heat shock protein that is a homolog of the Arabidopsis gene AtARC6, which is involved in chloroplast division. Knockdown and knockout of SlARC6 in wild-type tomato inhibit chloroplast division, indicating the conserved function of SlARC6. In su mutants, most mesophyll cells contain only one or two giant chloroplasts, while no chloroplasts are visible in 60% of stem cells, resulting in the albinic phenotype. Compared with mature tissues, the meristem of su mutants suggested that chloroplasts could partially divide in meristematic cells, suggesting the existence of an alternative mechanism in those dividing cells. Interestingly, the adaxial petiole cells of su mutants contain more chloroplasts than the abaxial cells. In addition, prolonged lighting can partially rescue the albinic phenotypes in su mutants, implying that light may promote SlACR6-independent chloroplast development. Our results verify the role of SlACR6 in chloroplast division in tomato and uncover the tissue-specific regulation of chloroplast development.
Conflict of interest statement
The authors declare no competing interests.
Figures






Similar articles
-
An Arabidopsis chloroplast-targeted Hsp101 homologue, APG6, has an essential role in chloroplast development as well as heat-stress response.Plant J. 2006 Oct;48(2):249-60. doi: 10.1111/j.1365-313X.2006.02873.x. Epub 2006 Sep 22. Plant J. 2006. PMID: 16995899
-
A Mutation in GIANT CHLOROPLAST Encoding a PARC6 Homolog Affects Spikelet Fertility in Rice.Plant Cell Physiol. 2015 May;56(5):977-91. doi: 10.1093/pcp/pcv024. Epub 2015 Feb 8. Plant Cell Physiol. 2015. PMID: 25667114
-
A Genetic Analysis of Chloroplast Division and Expansion in Arabidopsis thaliana.Plant Physiol. 1994 Jan;104(1):201-207. doi: 10.1104/pp.104.1.201. Plant Physiol. 1994. PMID: 12232072 Free PMC article.
-
Characterization of chloroplast division using the Arabidopsis mutant arc5.Plant Physiol. 1996 Sep;112(1):149-59. doi: 10.1104/pp.112.1.149. Plant Physiol. 1996. PMID: 8819323 Free PMC article.
-
Purine biosynthetic enzyme ATase2 is involved in the regulation of early chloroplast development and chloroplast gene expression in Arabidopsis.Photosynth Res. 2015 Dec;126(2-3):285-300. doi: 10.1007/s11120-015-0131-z. Epub 2015 Apr 3. Photosynth Res. 2015. PMID: 25837856
Cited by
-
Mutation of CsARC6 affects fruit color and increases fruit nutrition in cucumber.Theor Appl Genet. 2023 Apr 13;136(5):111. doi: 10.1007/s00122-023-04337-7. Theor Appl Genet. 2023. PMID: 37052704
-
Fine Mapping and Candidate Gene Analysis of BnC08.cds, a Recessive Gene Responsible for Sepal-Specific Chlorophyll-Deficiency in Brassica napus L.Front Plant Sci. 2022 Mar 10;13:850330. doi: 10.3389/fpls.2022.850330. eCollection 2022. Front Plant Sci. 2022. PMID: 35360306 Free PMC article.
-
Genome-wide identification of DnaJ gene family in Catalpa bungei and functional analysis of CbuDnaJ49 in leaf color formation.Front Plant Sci. 2023 Feb 23;14:1116063. doi: 10.3389/fpls.2023.1116063. eCollection 2023. Front Plant Sci. 2023. PMID: 36968394 Free PMC article.
-
Integrating physiology, genetics, and transcriptome to decipher a new thermo-sensitive and light-sensitive virescent leaf gene mutant in cucumber.Front Plant Sci. 2022 Aug 16;13:972620. doi: 10.3389/fpls.2022.972620. eCollection 2022. Front Plant Sci. 2022. PMID: 36051299 Free PMC article.
-
Genome-wide identification of GA2ox genes family and analysis of PbrGA2ox1-mediated enhanced chlorophyll accumulation by promoting chloroplast development in pear.BMC Plant Biol. 2024 Mar 4;24(1):166. doi: 10.1186/s12870-024-04842-x. BMC Plant Biol. 2024. PMID: 38433195 Free PMC article.
References
-
- Possingham JV, Lawrence ME. Controls to chloroplast division. Int. Rev. Cytol. 1983;84:1–56. doi: 10.1016/S0074-7696(08)61014-1. - DOI
