

High glucose induced c-Met activation promotes aggressive phenotype and regulates expression of glucose metabolism genes in HCC cells
- PMID: 34059694
- PMCID: PMC8166976
- DOI: 10.1038/s41598-021-89765-5
High glucose induced c-Met activation promotes aggressive phenotype and regulates expression of glucose metabolism genes in HCC cells
Abstract
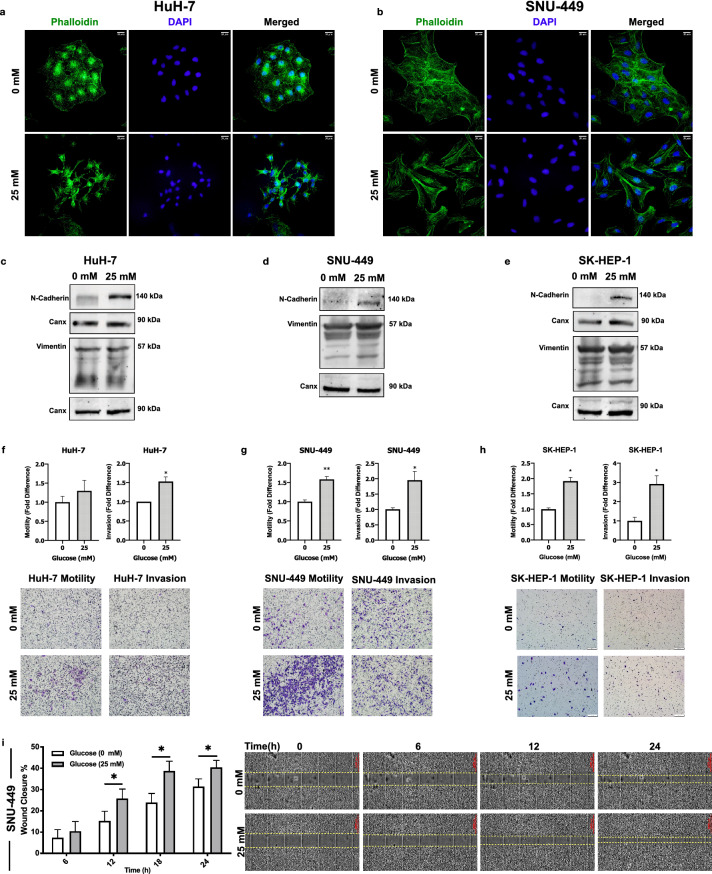
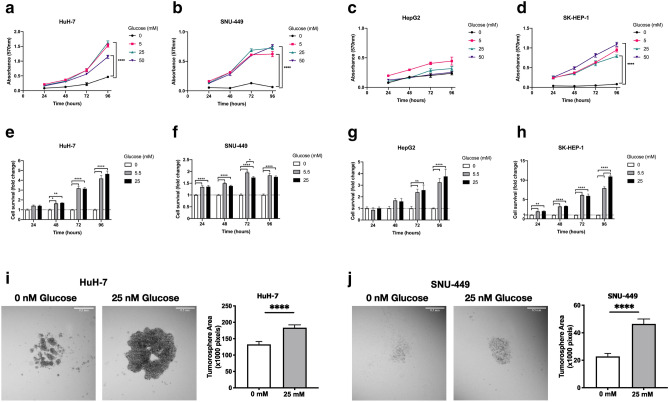
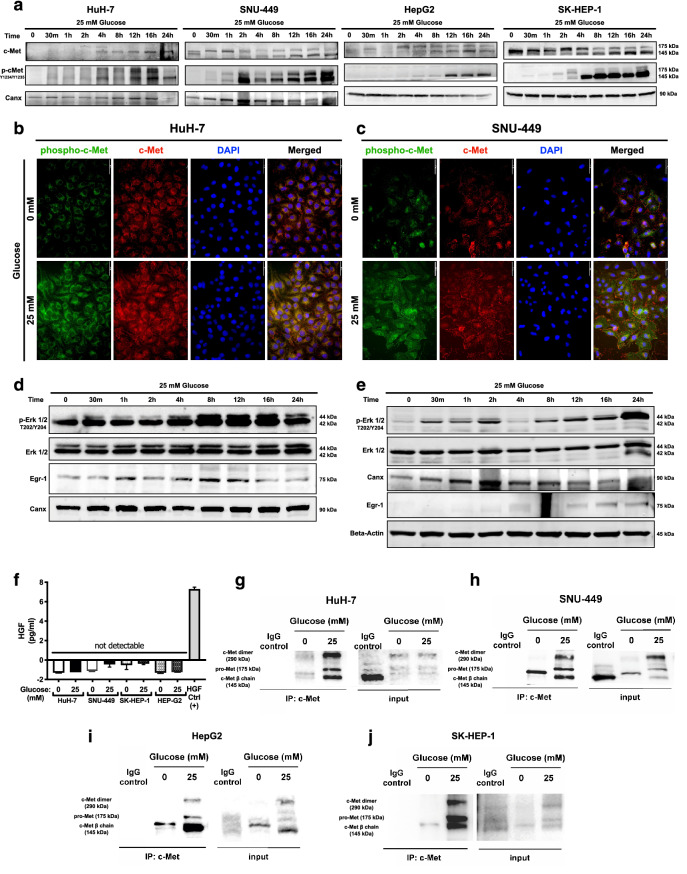
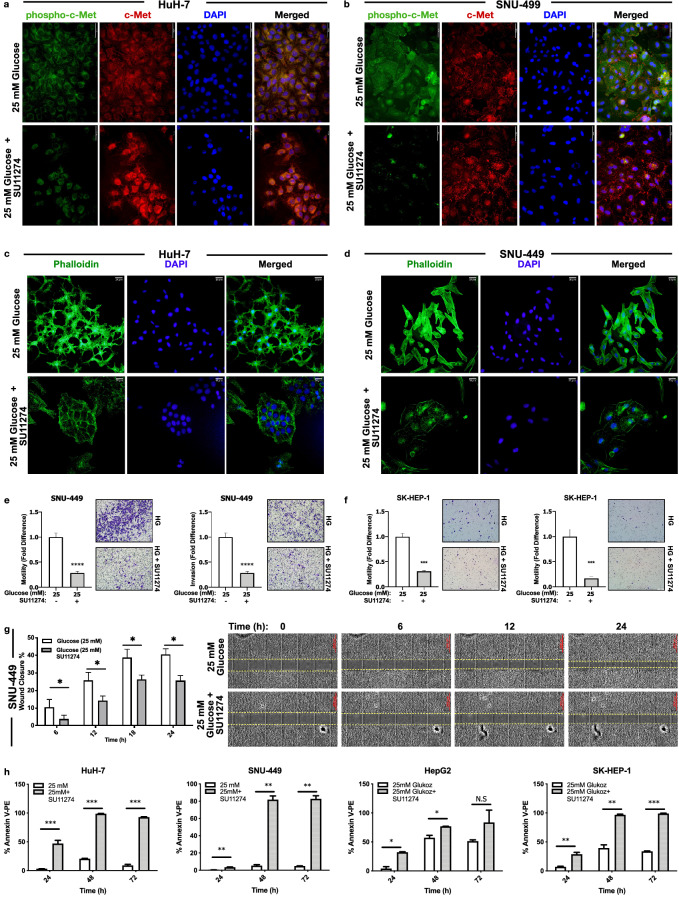
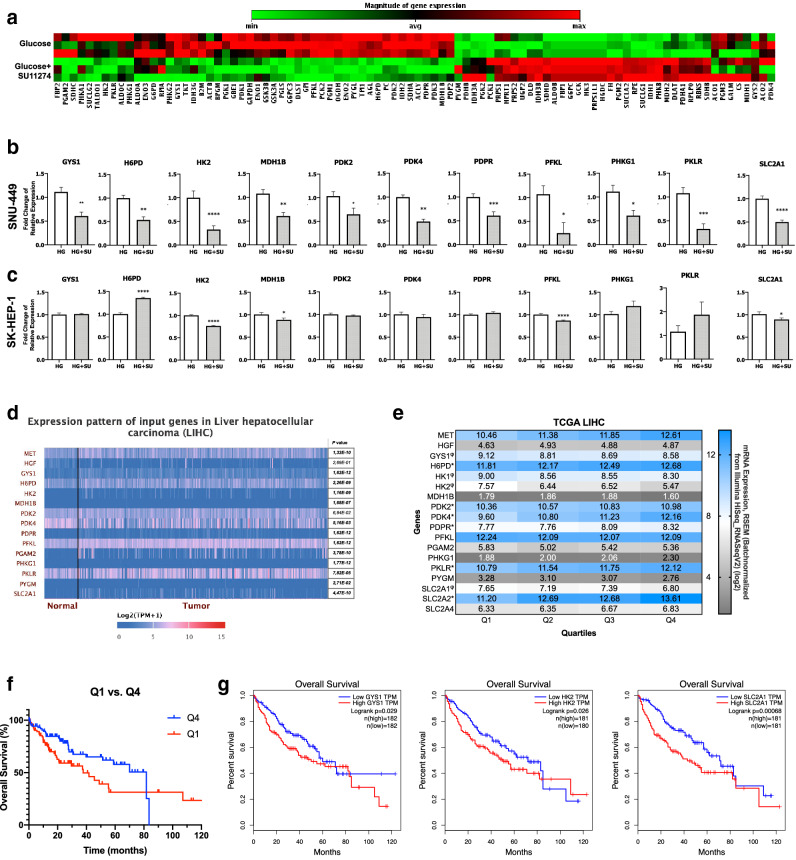
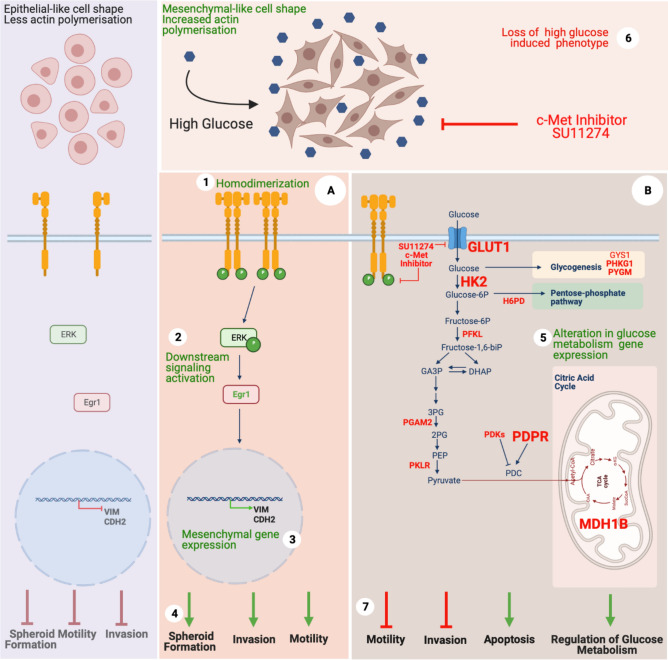
Hepatocellular carcinoma (HCC) is strongly associated with metabolic dysregulations/deregulations and hyperglycemia is a common metabolic disturbance in metabolic diseases. Hyperglycemia is defined to promote epithelial to mesenchymal transition (EMT) of cancer cells in various cancers but its molecular contribution to HCC progression and aggressiveness is relatively unclear. In this study, we analyzed the molecular mechanisms behind the hyperglycemia-induced EMT in HCC cell lines. Here, we report that high glucose promotes EMT through activating c-Met receptor tyrosine kinase via promoting its ligand-independent homodimerization. c-Met activation is critical for high glucose induced acquisition of mesenchymal phenotype, survival under high glucose stress and reprogramming of cellular metabolism by modulating glucose metabolism gene expression to promote aggressiveness in HCC cells. The crucial role of c-Met in high glucose induced EMT and aggressiveness may be the potential link between metabolic syndrome-related hepatocarcinogenesis and/or HCC progression. Considering c-Met inhibition in hyperglycemic patients would be an important complementary strategy for therapy that favors sensitization of HCC cells to therapeutics.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
