

Tsukushi is essential for the formation of the posterior semicircular canal that detects gait performance
- PMID: 34061311
- PMCID: PMC8642578
- DOI: 10.1007/s12079-021-00627-1
Tsukushi is essential for the formation of the posterior semicircular canal that detects gait performance
Abstract
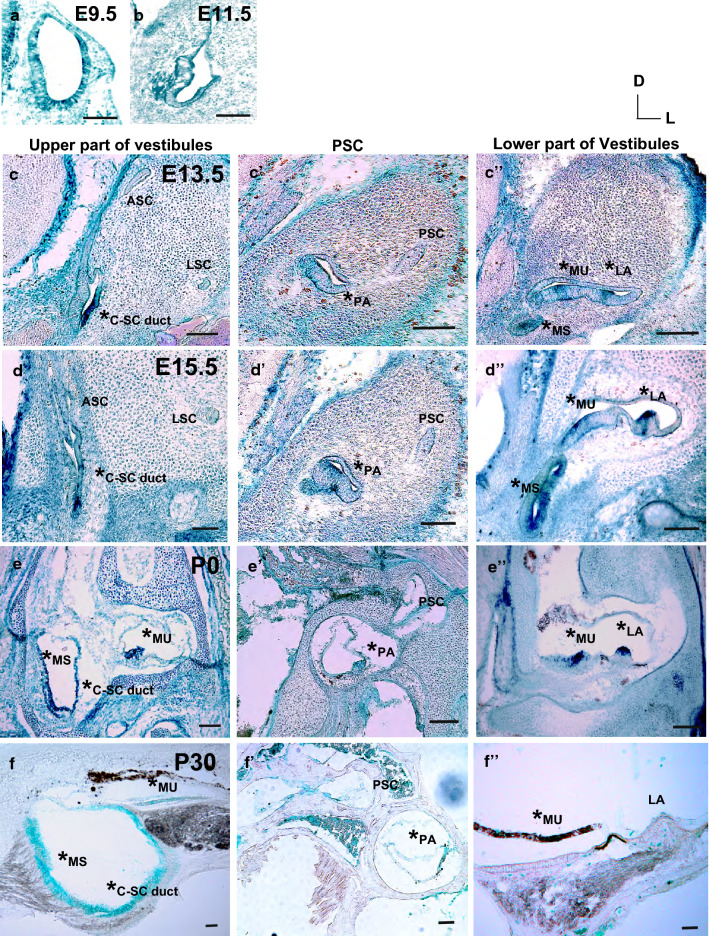
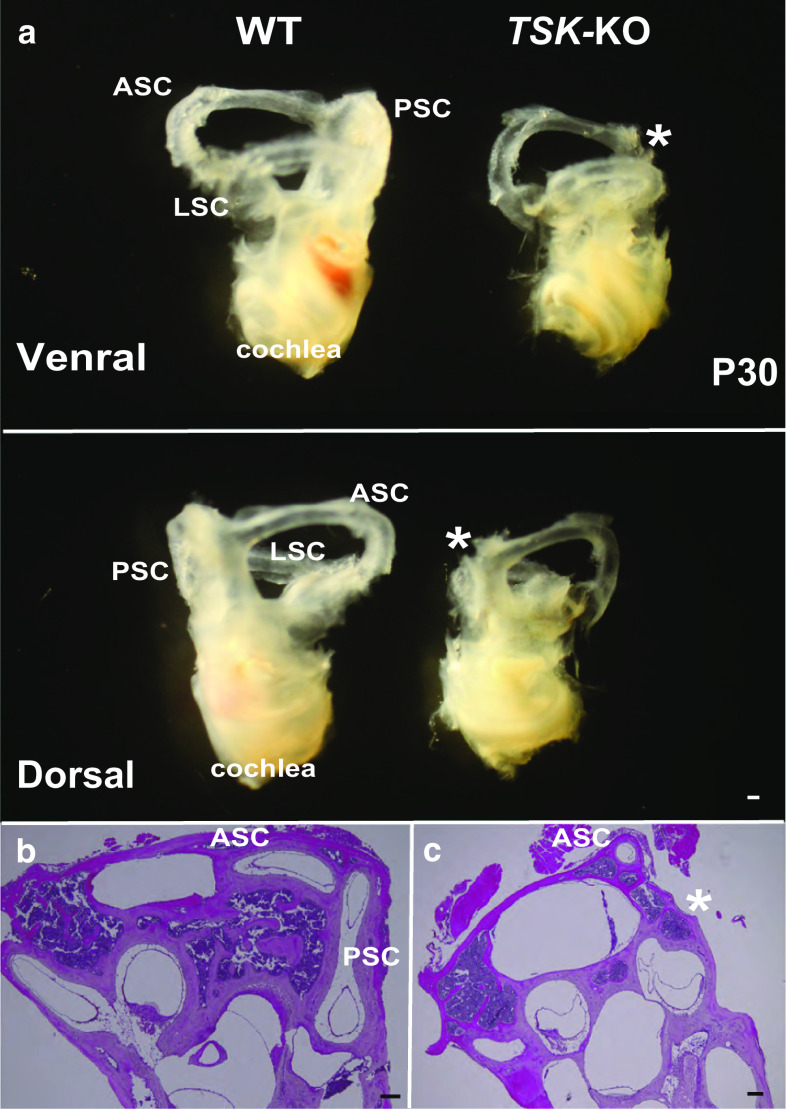
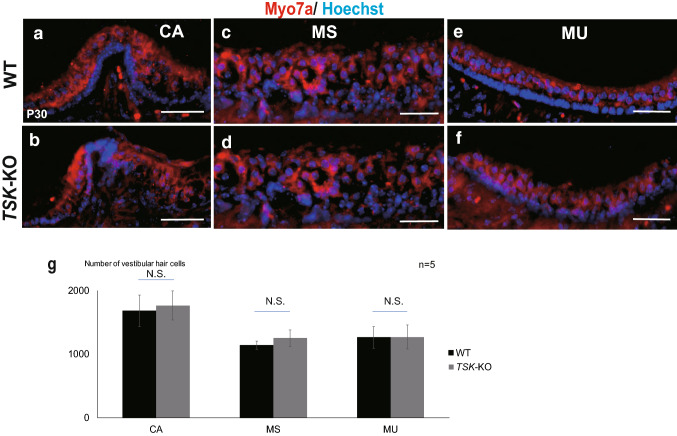
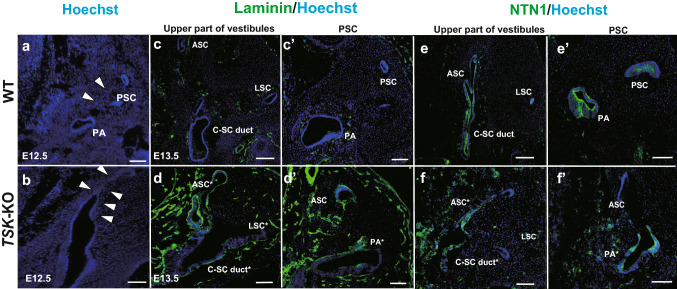
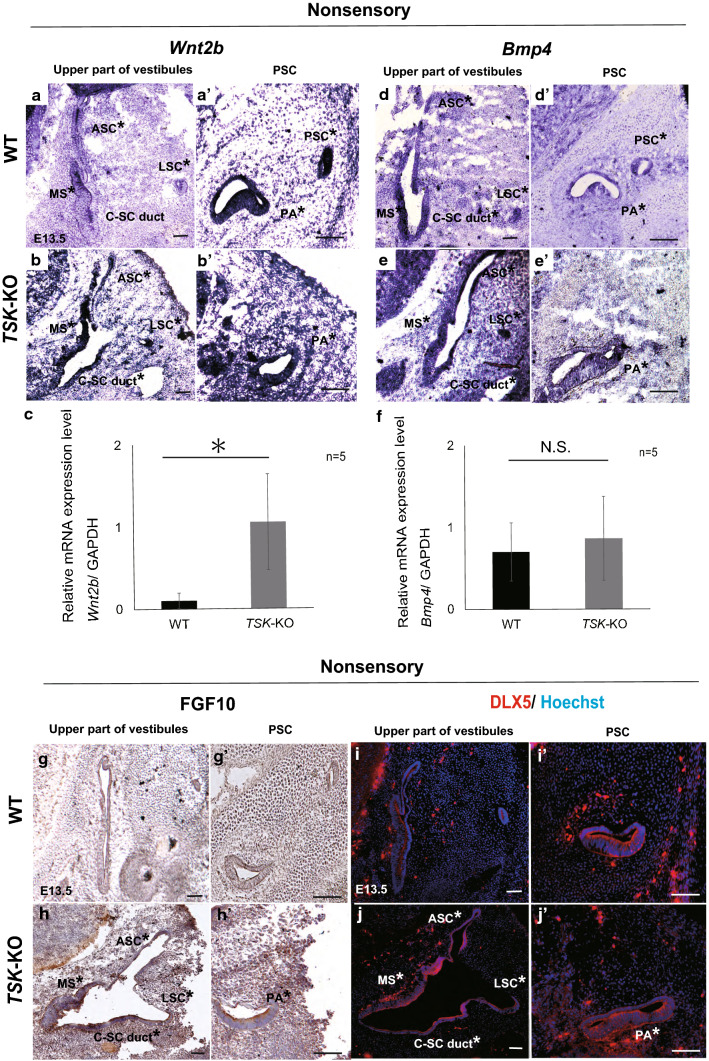
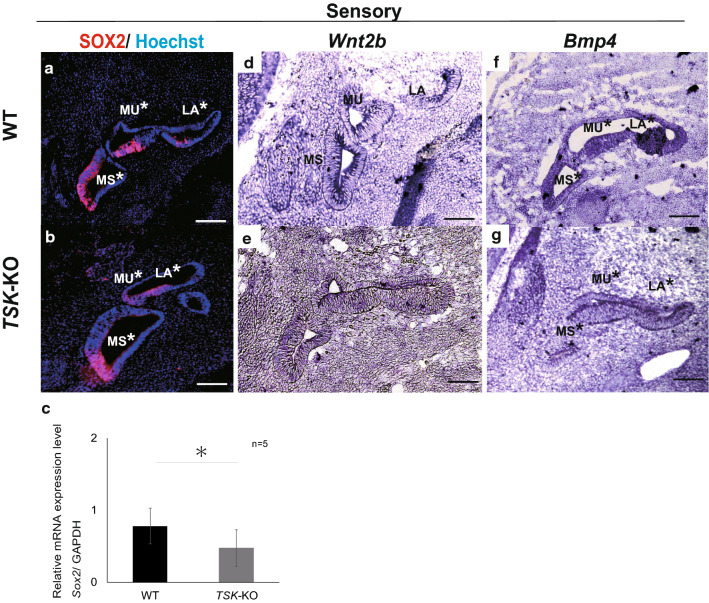
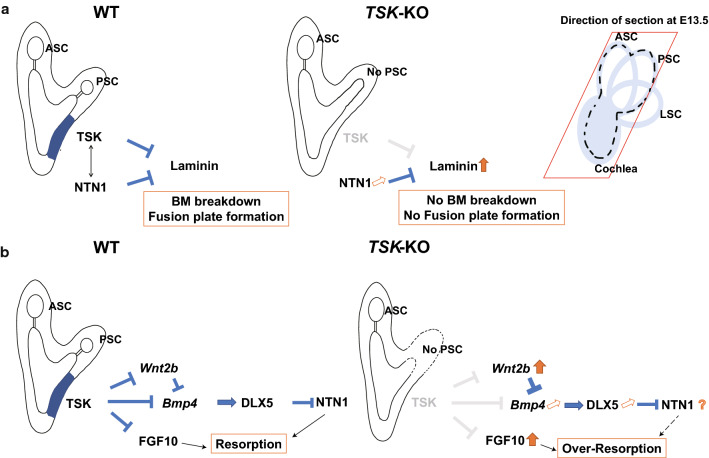
Tsukushi is a small, leucine-rich repeat proteoglycan that interacts with and regulates essential cellular signaling cascades in the chick retina and murine subventricular zone, hippocampus, dermal hair follicles, and the cochlea. However, its function in the vestibules of the inner ear remains unknown. Here, we investigated the function of Tsukushi in the vestibules and found that Tsukushi deficiency in mice resulted in defects in posterior semicircular canal formation in the vestibules, but did not lead to vestibular hair cell loss. Furthermore, Tsukushi accumulated in the non-prosensory and prosensory regions during the embryonic and postnatal developmental stages. The downregulation of Tsukushi altered the expression of key genes driving vestibule differentiation in the non-prosensory regions. Our results indicate that Tsukushi interacts with Wnt2b, bone morphogenetic protein 4, fibroblast growth factor 10, and netrin 1, thereby controlling semicircular canal formation. Therefore, Tsukushi may be an essential component of the molecular pathways regulating vestibular development.
Keywords: Bmp4; FGF10; Posterior semicircular canal; Tsukushi; Vestibule; Wnt2b.
© 2021. The International CCN Society.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
Similar articles
-
Tsukushi is essential for the development of the inner ear.Mol Brain. 2020 Mar 3;13(1):29. doi: 10.1186/s13041-020-00570-z. Mol Brain. 2020. PMID: 32127020 Free PMC article.
-
Bmp4 is essential for the formation of the vestibular apparatus that detects angular head movements.PLoS Genet. 2008 Apr 11;4(4):e1000050. doi: 10.1371/journal.pgen.1000050. PLoS Genet. 2008. PMID: 18404215 Free PMC article.
-
Fgf10 is required for specification of non-sensory regions of the cochlear epithelium.Dev Biol. 2015 Apr 1;400(1):59-71. doi: 10.1016/j.ydbio.2015.01.015. Epub 2015 Jan 24. Dev Biol. 2015. PMID: 25624266 Free PMC article.
-
Canal dehiscence.Curr Opin Neurol. 2011 Feb;24(1):25-31. doi: 10.1097/WCO.0b013e328341ef88. Curr Opin Neurol. 2011. PMID: 21124219 Review.
-
Anatomical and physiological development of the human inner ear.Hear Res. 2016 Aug;338:9-21. doi: 10.1016/j.heares.2016.02.004. Epub 2016 Feb 18. Hear Res. 2016. PMID: 26900072 Review.
Cited by
-
Association of serum Tsukushi level with metabolic syndrome and its components.Endocrine. 2023 Mar;79(3):469-476. doi: 10.1007/s12020-022-03285-4. Epub 2023 Jan 2. Endocrine. 2023. PMID: 36592295
-
Evolutionary analysis of genes associated with the sense of balance in semi-aquatic mammals.BMC Ecol Evol. 2025 Jan 10;25(1):8. doi: 10.1186/s12862-024-02345-9. BMC Ecol Evol. 2025. PMID: 39794719 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
