

A Peek into the Plasmidome of Global Sewage
- PMID: 34061588
- PMCID: PMC8269221
- DOI: 10.1128/mSystems.00283-21
A Peek into the Plasmidome of Global Sewage
Abstract
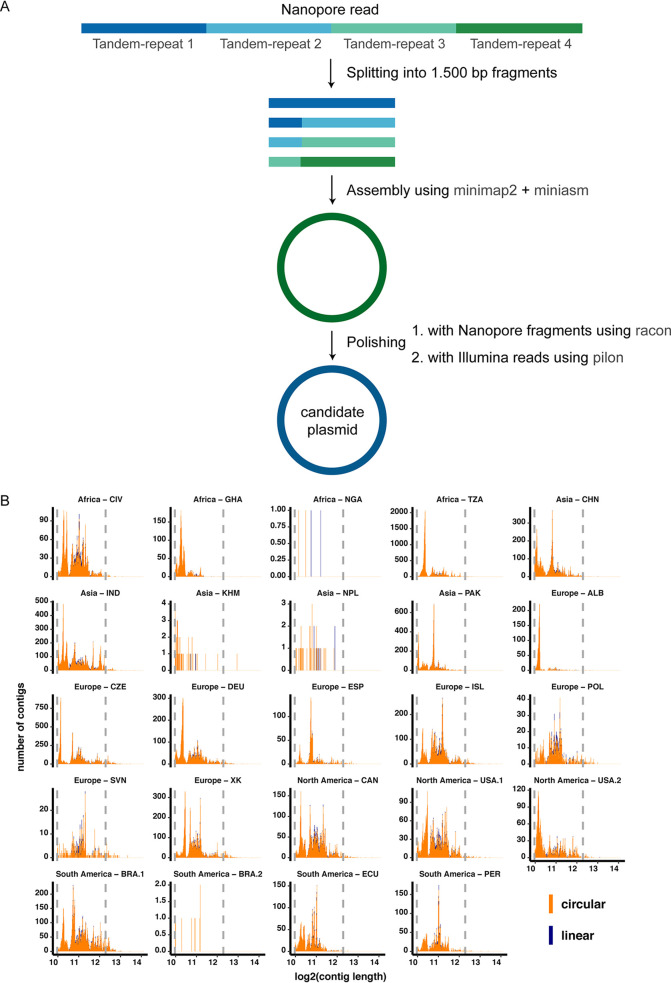
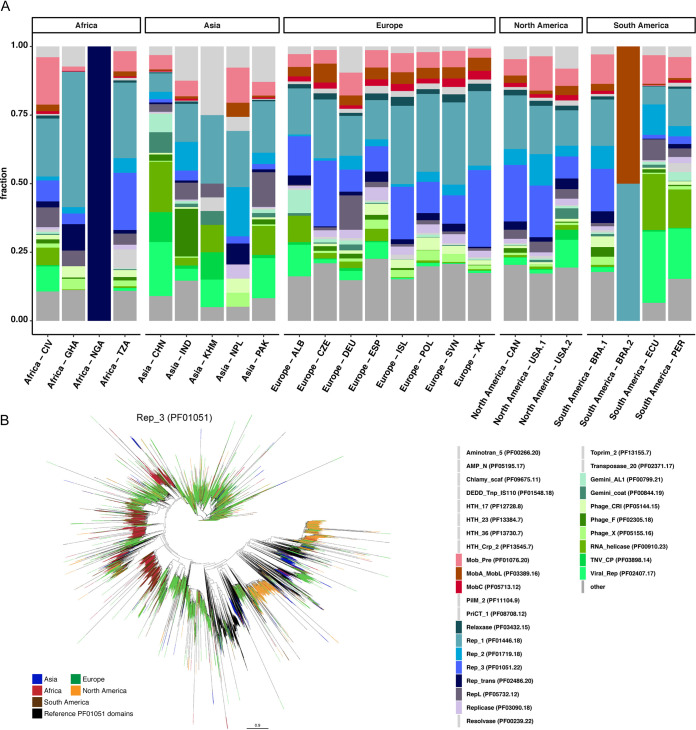
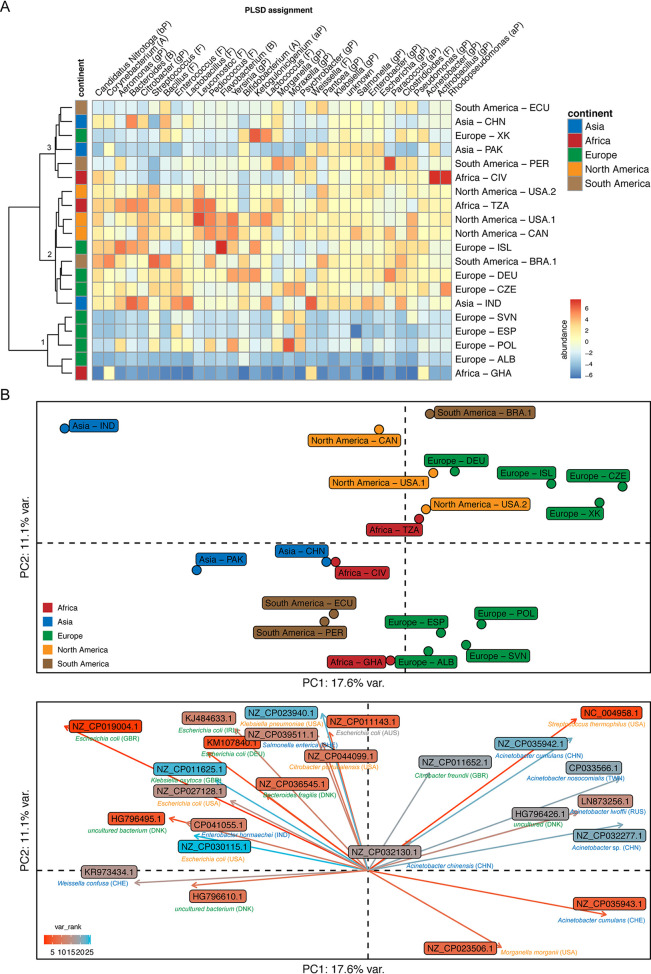
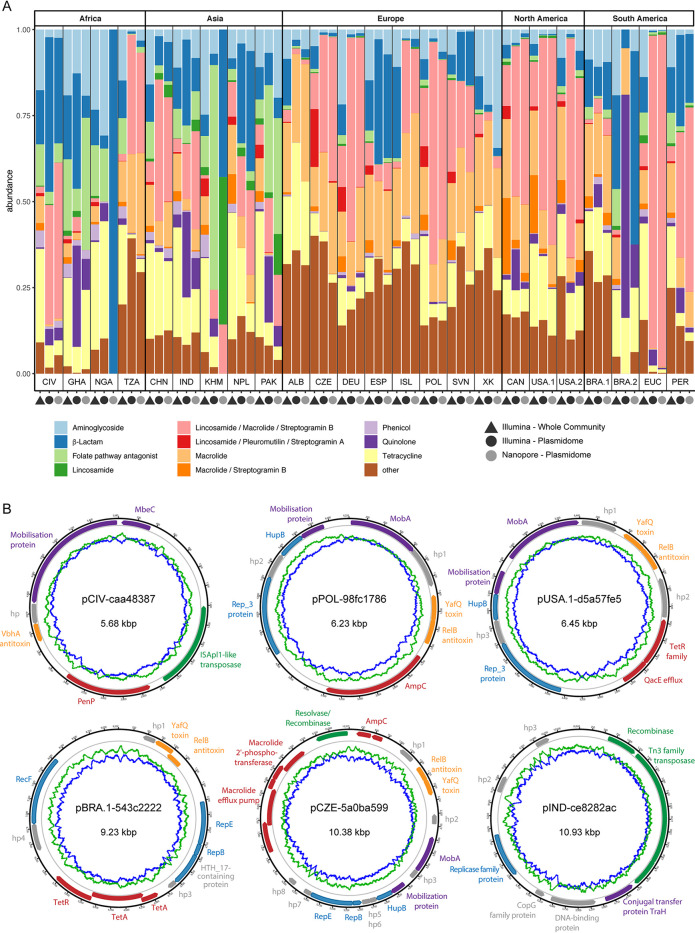
Plasmids can provide a selective advantage for microorganisms to survive and adapt to new environmental conditions. Plasmid-encoded traits, such as antimicrobial resistance (AMR) or virulence, impact the ecology and evolution of bacteria and can significantly influence the burden of infectious diseases. Insight about the identity and functions encoded on plasmids on the global scale are largely lacking. Here, we investigate the plasmidome of 24 samples (22 countries, 5 continents) from the global sewage surveillance project. We obtained 105-Gbp Oxford Nanopore and 167-Gbp Illumina NextSeq DNA sequences from plasmid DNA preparations and assembled 165,302 contigs (159,322 circular). Of these, 58,429 carried genes encoding for plasmid-related and 11,222 for virus/phage-related proteins. About 90% of the circular DNA elements did not have any similarity to known plasmids. Those that exhibited similarity had similarity to plasmids whose hosts were previously detected in these sewage samples (e.g., Acinetobacter, Escherichia, Moraxella, Enterobacter, Bacteroides, and Klebsiella). Some AMR classes were detected at a higher abundance in plasmidomes (e.g., macrolide-lincosamide-streptogramin B, macrolide, and quinolone) compared to the respective complex sewage samples. In addition to AMR genes, a range of functions were encoded on the candidate plasmids, including plasmid replication and maintenance, mobilization, and conjugation. In summary, we describe a laboratory and bioinformatics workflow for the recovery of plasmids and other potential extrachromosomal DNA elements from complex microbiomes. Moreover, the obtained data could provide further valuable insight into the ecology and evolution of microbiomes, knowledge about AMR transmission, and the discovery of novel functions. IMPORTANCE This is, to the best of our knowledge, the first study to investigate plasmidomes at a global scale using long read sequencing from complex untreated domestic sewage. Previous metagenomic surveys have detected AMR genes in a variety of environments, including sewage. However, it is unknown whether the AMR genes were present on the microbial chromosome or located on extrachromosomal elements, such as plasmids. Using our approach, we recovered a large number of plasmids, of which most appear novel. We identified distinct AMR genes that were preferentially located on plasmids, potentially contributing to their transmissibility. Overall, plasmids are of great importance for the biology of microorganisms in their natural environments (free-living and host-associated), as well as for molecular biology and biotechnology. Plasmidome collections may therefore be valuable resources for the discovery of fundamental biological mechanisms and novel functions useful in a variety of contexts.
Keywords: Illumina NextSeq; Oxford Nanopore; Oxford Nanopore sequencing; animals; human; microbiome; plasmid DNA isolation; plasmid reconstruction; plasmids; wastewater.
Figures
Similar articles
-
Global Distribution and Diversity of Prevalent Sewage Water Plasmidomes.mSystems. 2022 Oct 26;7(5):e0019122. doi: 10.1128/msystems.00191-22. Epub 2022 Sep 7. mSystems. 2022. PMID: 36069451 Free PMC article.
-
Large Circular Plasmids from Groundwater Plasmidomes Span Multiple Incompatibility Groups and Are Enriched in Multimetal Resistance Genes.mBio. 2019 Feb 26;10(1):e02899-18. doi: 10.1128/mBio.02899-18. mBio. 2019. PMID: 30808697 Free PMC article.
-
Plasmids Shaped the Recent Emergence of the Major Nosocomial Pathogen Enterococcus faecium.mBio. 2020 Feb 11;11(1):e03284-19. doi: 10.1128/mBio.03284-19. mBio. 2020. PMID: 32047136 Free PMC article.
-
Metamobilomics--expanding our knowledge on the pool of plasmid encoded traits in natural environments using high-throughput sequencing.Clin Microbiol Infect. 2012 Jul;18 Suppl 4:5-7. doi: 10.1111/j.1469-0691.2012.03862.x. Clin Microbiol Infect. 2012. PMID: 22647039 Review.
-
Bacterial plasmids: replication of extrachromosomal genetic elements encoding resistance to antimicrobial compounds.Front Biosci. 1999 Jan 1;4:D43-62. doi: 10.2741/actis. Front Biosci. 1999. PMID: 9872728 Review.
Cited by
-
Comprehensive Genome and Plasmidome Analysis of Antimicrobial Resistant Bacteria in Wastewater Treatment Plant Effluent of Tokyo.Antibiotics (Basel). 2022 Sep 21;11(10):1283. doi: 10.3390/antibiotics11101283. Antibiotics (Basel). 2022. PMID: 36289941 Free PMC article.
-
Integrative genomics would strengthen AMR understanding through ONE health approach.Heliyon. 2024 Jul 17;10(14):e34719. doi: 10.1016/j.heliyon.2024.e34719. eCollection 2024 Jul 30. Heliyon. 2024. PMID: 39816336 Free PMC article. Review.
-
Global Distribution and Diversity of Prevalent Sewage Water Plasmidomes.mSystems. 2022 Oct 26;7(5):e0019122. doi: 10.1128/msystems.00191-22. Epub 2022 Sep 7. mSystems. 2022. PMID: 36069451 Free PMC article.
-
Characterization of Carbapenem-Resistant Enterobacteriaceae Cultured From Retail Meat Products, Patients, and Porcine Excrement in China.Front Microbiol. 2021 Dec 23;12:743468. doi: 10.3389/fmicb.2021.743468. eCollection 2021. Front Microbiol. 2021. PMID: 35002997 Free PMC article.
-
Mixed waste contamination selects for a mobile genetic element population enriched in multiple heavy metal resistance genes.ISME Commun. 2024 May 9;4(1):ycae064. doi: 10.1093/ismeco/ycae064. eCollection 2024 Jan. ISME Commun. 2024. PMID: 38800128 Free PMC article.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
