

Transient Mitochondria Dysfunction Confers Fungal Cross-Resistance against Phagocytic Killing and Fluconazole
- PMID: 34061590
- PMCID: PMC8262853
- DOI: 10.1128/mBio.01128-21
Transient Mitochondria Dysfunction Confers Fungal Cross-Resistance against Phagocytic Killing and Fluconazole
Abstract
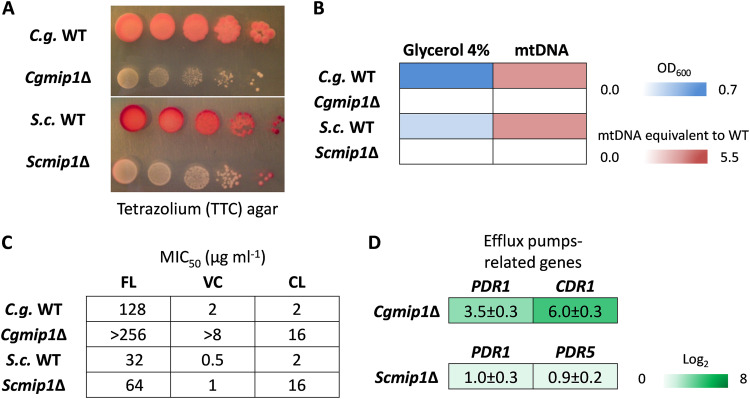
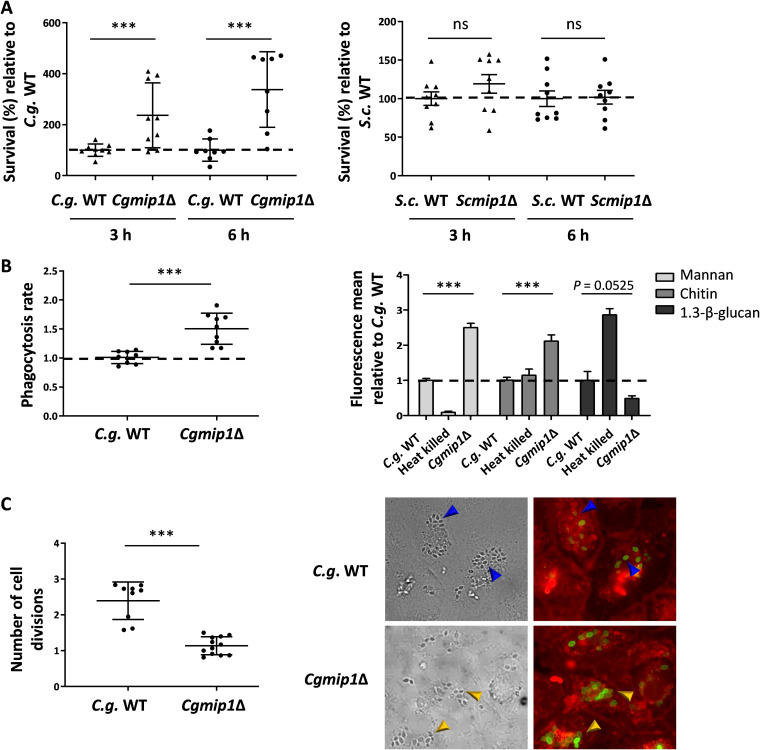
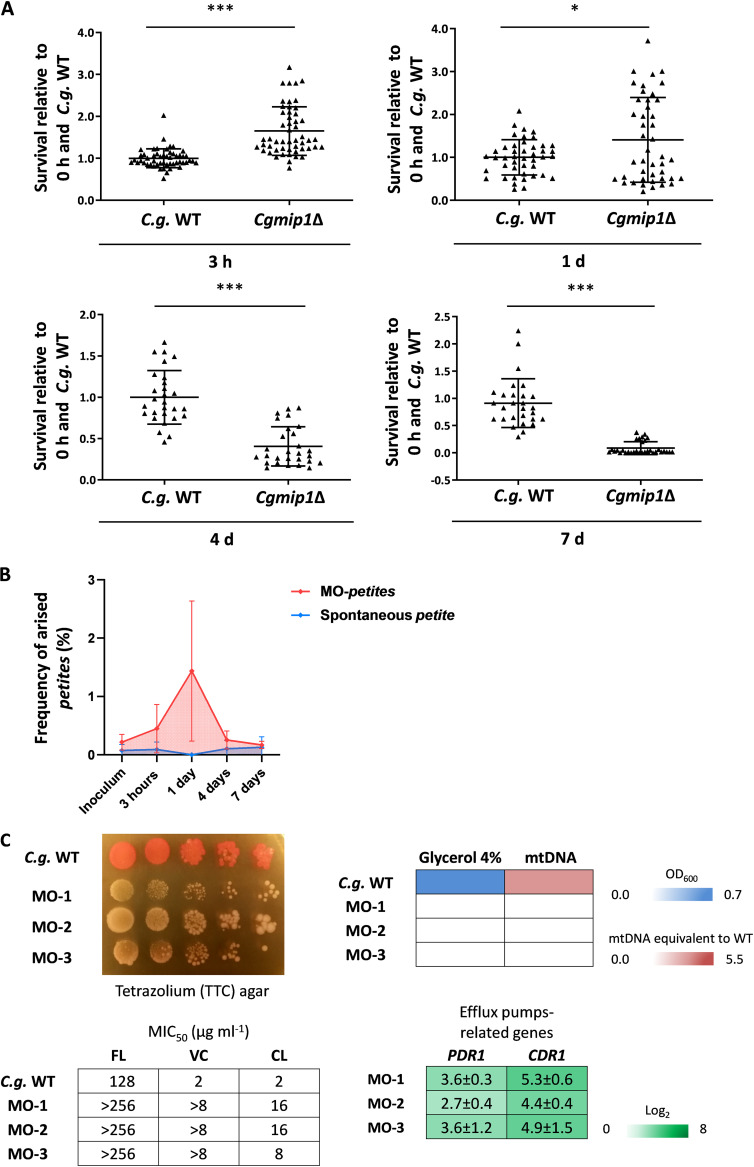
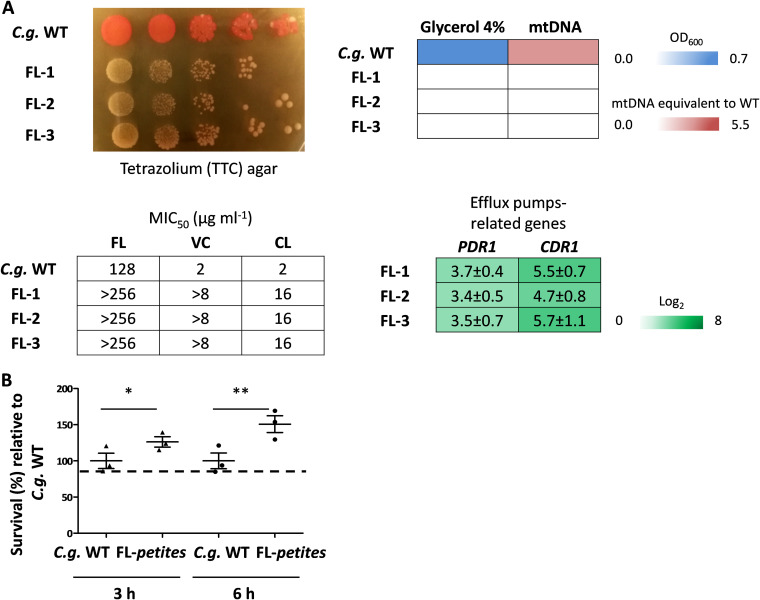
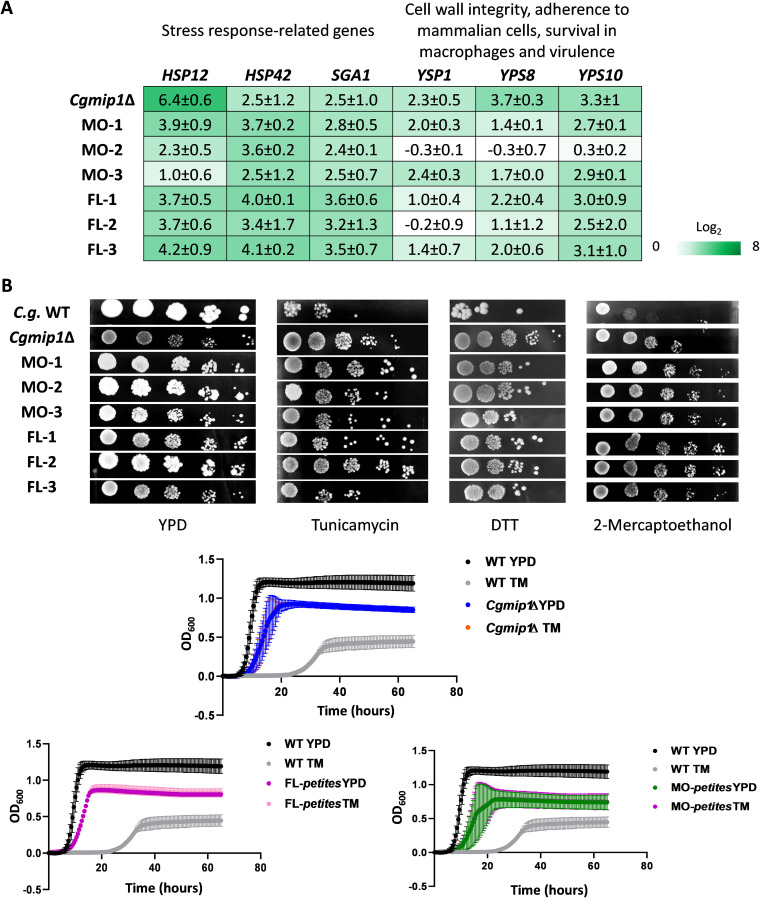
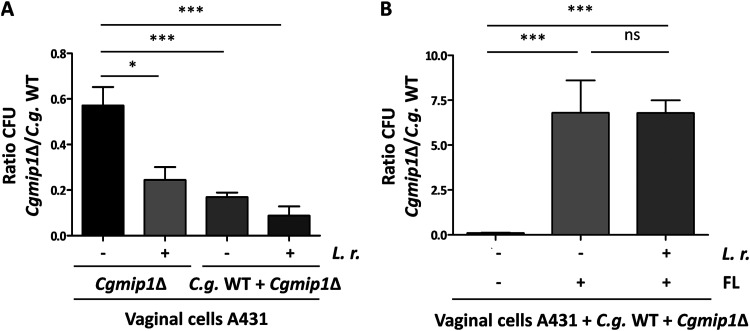
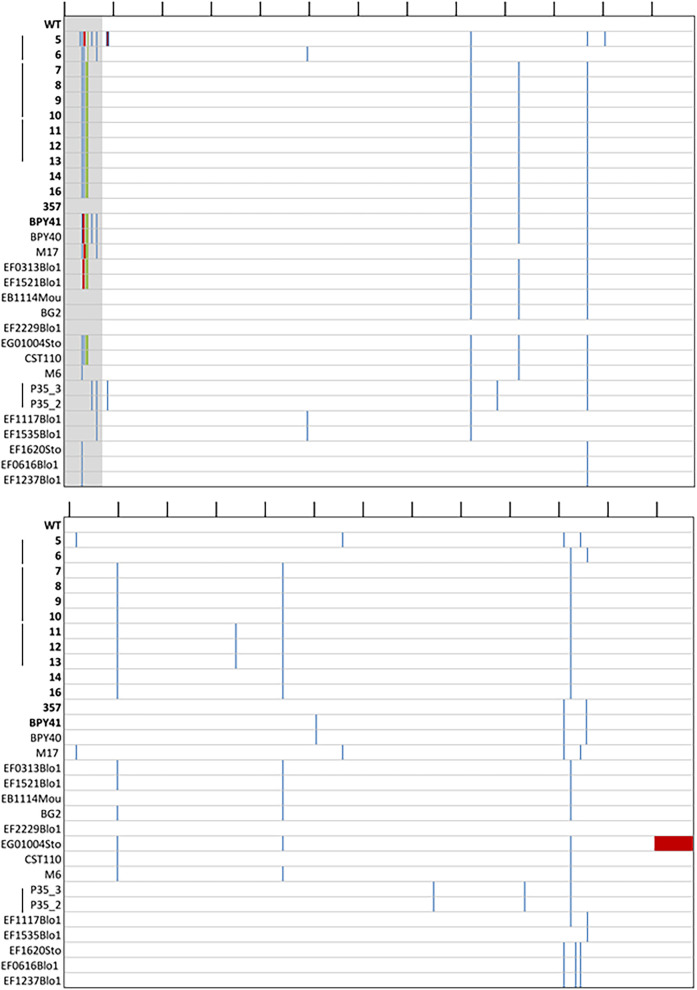
Loss or inactivation of antivirulence genes is an adaptive strategy in pathogen evolution. Candida glabrata is an important opportunistic pathogen related to baker's yeast, with the ability to both quickly increase its intrinsic high level of azole resistance and persist within phagocytes. During C. glabrata's evolution as a pathogen, the mitochondrial DNA polymerase CgMip1 has been under positive selection. We show that CgMIP1 deletion not only triggers loss of mitochondrial function and a petite phenotype, but increases C. glabrata's azole and endoplasmic reticulum (ER) stress resistance and, importantly, its survival in phagocytes. The same phenotype is induced by fluconazole and by exposure to macrophages, conferring a cross-resistance between antifungals and immune cells, and can be found in clinical isolates despite a slow growth of petite strains. This suggests that petite constitutes a bet-hedging strategy of C. glabrata and, potentially, a relevant cause of azole resistance. Mitochondrial function may therefore be considered a potential antivirulence factor. IMPORTANCE Candida glabrata is an opportunistic pathogen whose incidence has been increasing in the last 40 years. It has risen to become the most prominent non-Candida albicans Candida (NCAC) species to cause candidemia, constituting about one-third of isolates in the United States, and steadily increasing in European countries and in Australia. Despite its clinical importance, C. glabrata's pathogenicity strategies remain poorly understood. Our research shows that loss of mitochondrial function and the resulting petite phenotype is advantageous for C. glabrata to cope with infection-related stressors, such as antifungals and host immune defenses. The (cross-)resistance against both these factors may have major implications in the clinical outcome of infections with this major fungal pathogen.
Keywords: antivirulence; cross-resistance; fungal infection; petite.
Figures
Similar articles
-
Temporal transcriptional response of Candida glabrata during macrophage infection reveals a multifaceted transcriptional regulator CgXbp1 important for macrophage response and fluconazole resistance.Elife. 2024 Oct 2;13:e73832. doi: 10.7554/eLife.73832. Elife. 2024. PMID: 39356739 Free PMC article.
-
Contribution of CgPDR1-regulated genes in enhanced virulence of azole-resistant Candida glabrata.PLoS One. 2011 Mar 9;6(3):e17589. doi: 10.1371/journal.pone.0017589. PLoS One. 2011. PMID: 21408004 Free PMC article.
-
Mechanisms of azole resistance in clinical isolates of Candida glabrata collected during a hospital survey of antifungal resistance.Antimicrob Agents Chemother. 2005 Feb;49(2):668-79. doi: 10.1128/AAC.49.2.668-679.2005. Antimicrob Agents Chemother. 2005. PMID: 15673750 Free PMC article.
-
Candida and candidaemia. Susceptibility and epidemiology.Dan Med J. 2013 Nov;60(11):B4698. Dan Med J. 2013. PMID: 24192246 Review.
-
Resistance in human pathogenic yeasts and filamentous fungi: prevalence, underlying molecular mechanisms and link to the use of antifungals in humans and the environment.Dan Med J. 2016 Oct;63(10):B5288. Dan Med J. 2016. PMID: 27697142 Review.
Cited by
-
Candida albicans and Candida glabrata: global priority pathogens.Microbiol Mol Biol Rev. 2024 Jun 27;88(2):e0002123. doi: 10.1128/mmbr.00021-23. Epub 2024 Jun 4. Microbiol Mol Biol Rev. 2024. PMID: 38832801 Free PMC article. Review.
-
Disclosing azole resistance mechanisms in resistant Candida glabrata strains encoding wild-type or gain-of-function CgPDR1 alleles through comparative genomics and transcriptomics.G3 (Bethesda). 2022 Jul 6;12(7):jkac110. doi: 10.1093/g3journal/jkac110. G3 (Bethesda). 2022. PMID: 35532173 Free PMC article.
-
Overlooked Candida glabrata petites are echinocandin tolerant, induce host inflammatory responses, and display poor in vivo fitness.bioRxiv [Preprint]. 2023 Jun 16:2023.06.15.545195. doi: 10.1101/2023.06.15.545195. bioRxiv. 2023. Update in: mBio. 2023 Oct 31;14(5):e0118023. doi: 10.1128/mbio.01180-23. PMID: 37398397 Free PMC article. Updated. Preprint.
-
Genotypic diversity and unrecognized antifungal resistance among populations of Candida glabrata from positive blood cultures.Res Sq [Preprint]. 2023 Apr 3:rs.3.rs-2706400. doi: 10.21203/rs.3.rs-2706400/v1. Res Sq. 2023. Update in: Nat Commun. 2023 Sep 22;14(1):5918. doi: 10.1038/s41467-023-41509-x. PMID: 37066226 Free PMC article. Updated. Preprint.
-
Uncovering the connection between tunicamycin-induced respiratory deficiency and reduced fluconazole tolerance in Candida glabrata.Front Microbiol. 2025 Apr 28;16:1528341. doi: 10.3389/fmicb.2025.1528341. eCollection 2025. Front Microbiol. 2025. PMID: 40356653 Free PMC article.
References
-
- Gabaldon T, Martin T, Marcet-Houben M, Durrens P, Bolotin-Fukuhara M, Lespinet O, Arnaise S, Boisnard S, Aguileta G, Atanasova R, Bouchier C, Couloux A, Creno S, Cruz JA, Devillers H, Enache-Angoulvant A, Guitard J, Jaouen L, Ma L, Marck C, Neuvéglise C, Pelletier E, Pinard A, Poulain J, Recoquillay J, Westhof E, Wincker P, Dujon B, Hennequin C, Fairhead C. 2013. Comparative genomics of emerging pathogens in the Candida glabrata clade. BMC Genomics 14:623. 10.1186/1471-2164-14-623. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
