

Predicting direct and indirect non-target impacts of biocontrol agents using machine-learning approaches
- PMID: 34061885
- PMCID: PMC8168882
- DOI: 10.1371/journal.pone.0252448
Predicting direct and indirect non-target impacts of biocontrol agents using machine-learning approaches
Erratum in
-
Correction: Predicting direct and indirect non-target impacts of biocontrol agents using machine-learning approaches.PLoS One. 2021 Sep 29;16(9):e0258080. doi: 10.1371/journal.pone.0258080. eCollection 2021. PLoS One. 2021. PMID: 34587224 Free PMC article.
Abstract
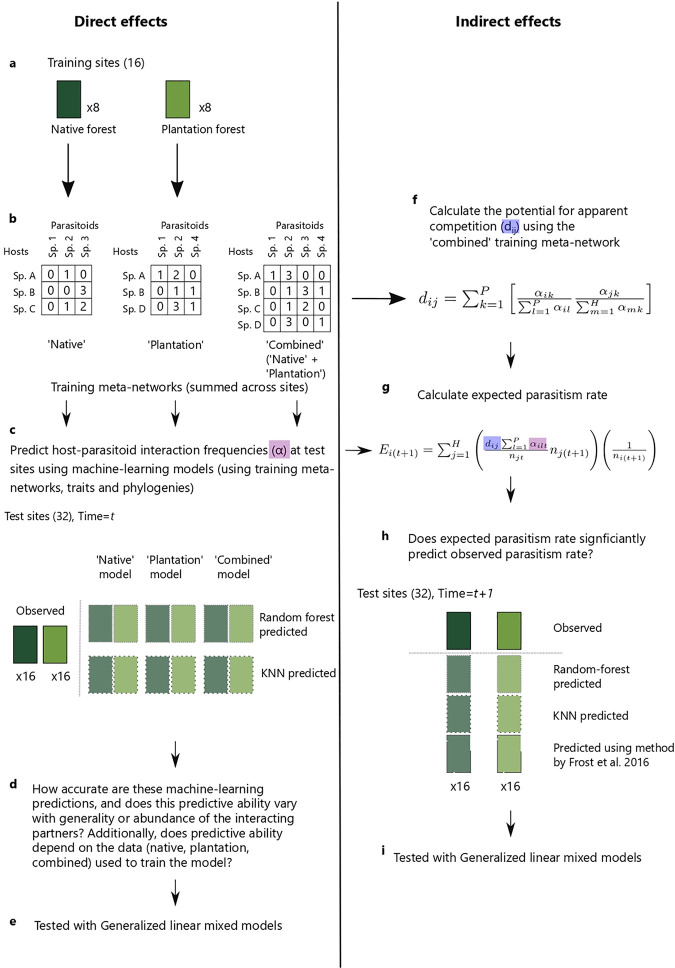
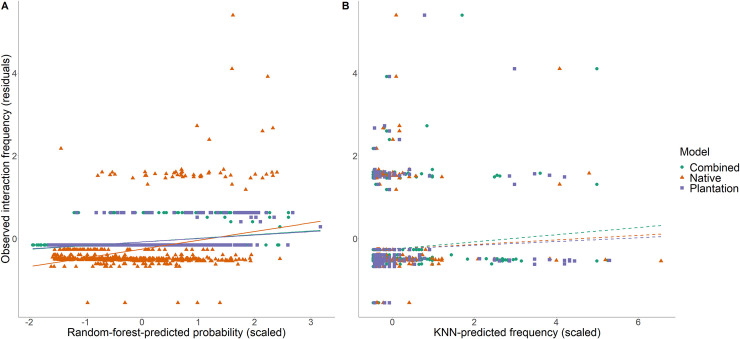
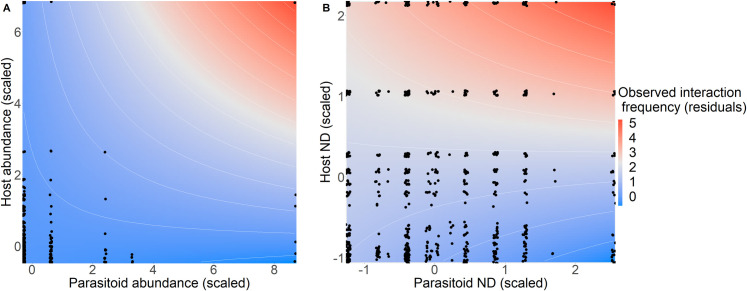
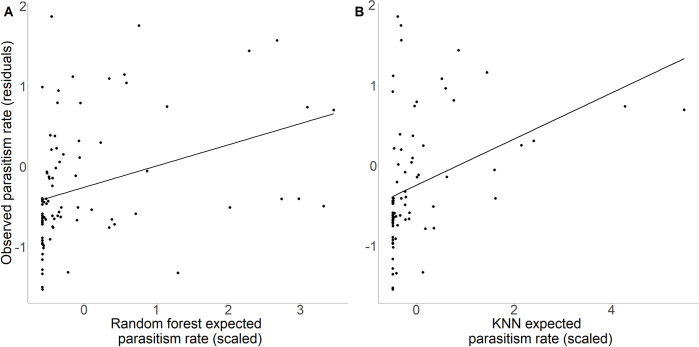
Biological pest control (i.e. 'biocontrol') agents can have direct and indirect non-target impacts, and predicting these effects (especially indirect impacts) remains a central challenge in biocontrol risk assessment. The analysis of ecological networks offers a promising approach to understanding the community-wide impacts of biocontrol agents (via direct and indirect interactions). Independently, species traits and phylogenies have been shown to successfully predict species interactions and network structure (alleviating the need to collect quantitative interaction data), but whether these approaches can be combined to predict indirect impacts of natural enemies remains untested. Whether predictions of interactions (i.e. direct effects) can be made equally well for generalists vs. specialists, abundant vs. less abundant species, and across different habitat types is also untested for consumer-prey interactions. Here, we used two machine-learning techniques (random forest and k-nearest neighbour; KNN) to test whether we could accurately predict empirically-observed quantitative host-parasitoid networks using trait and phylogenetic information. Then, we tested whether the accuracy of machine-learning-predicted interactions depended on the generality or abundance of the interacting partners, or on the source (habitat type) of the training data. Finally, we used these predicted networks to generate predictions of indirect effects via shared natural enemies (i.e. apparent competition), and tested these predictions against empirically observed indirect effects between hosts. We found that random-forest models predicted host-parasitoid pairwise interactions (which could be used to predict attack of non-target host species) more successfully than KNN. This predictive ability depended on the generality of the interacting partners for KNN models, and depended on species' abundances for both random-forest and KNN models, but did not depend on the source (habitat type) of data used to train the models. Further, although our machine-learning informed methods could significantly predict indirect effects, the explanatory power of our machine-learning models for indirect interactions was reasonably low. Combining machine-learning and network approaches provides a starting point for reducing risk in biocontrol introductions, and could be applied more generally to predicting species interactions such as impacts of invasive species.
Conflict of interest statement
The authors have declared that no competing interests exist. We acknowledge that one of the authors (J.H.T) is employed by a commercial company (The New Zealand Institute for Plant and Food Research Limited). This does not alter our adherence to PLOS ONE policies on sharing data and materials.
Figures
Similar articles
-
Four-trophic level food webs reveal the cascading impacts of an invasive plant targeted for biocontrol.Ecology. 2017 Mar;98(3):782-793. doi: 10.1002/ecy.1701. Ecology. 2017. PMID: 27987302
-
Molecular detection of trophic links in a complex insect host-parasitoid food web.Mol Ecol Resour. 2011 Sep;11(5):786-94. doi: 10.1111/j.1755-0998.2011.03016.x. Epub 2011 Apr 29. Mol Ecol Resour. 2011. PMID: 21535428
-
Ecological interactions and the Netflix problem.PeerJ. 2017 Aug 10;5:e3644. doi: 10.7717/peerj.3644. eCollection 2017. PeerJ. 2017. PMID: 28828250 Free PMC article.
-
Classical biological control of an invasive forest pest: a world perspective of the management of Sirex noctilio using the parasitoid Ibalia leucospoides (Hymenoptera: Ibaliidae).Bull Entomol Res. 2015 Feb;105(1):1-12. doi: 10.1017/S0007485314000418. Epub 2014 Jun 13. Bull Entomol Res. 2015. PMID: 24923367 Review.
-
Predicting non-target impacts.Curr Opin Insect Sci. 2020 Apr;38:79-83. doi: 10.1016/j.cois.2020.02.002. Epub 2020 Feb 27. Curr Opin Insect Sci. 2020. PMID: 32240966 Review.
Cited by
-
Correction: Predicting direct and indirect non-target impacts of biocontrol agents using machine-learning approaches.PLoS One. 2021 Sep 29;16(9):e0258080. doi: 10.1371/journal.pone.0258080. eCollection 2021. PLoS One. 2021. PMID: 34587224 Free PMC article.
-
Groundbreaking Technologies and the Biocontrol of Fungal Vascular Plant Pathogens.J Fungi (Basel). 2025 Jan 18;11(1):77. doi: 10.3390/jof11010077. J Fungi (Basel). 2025. PMID: 39852495 Free PMC article. Review.
References
-
- Pimentel D, Acquay H, Biltonen M, Rice P, Silva M, Nelson J, et al.. Environmental and economic costs of pesticide use. Bioscience. 1992;42(10):750–60.
Publication types
MeSH terms
Associated data
LinkOut - more resources
Full Text Sources
