

Complement Proteins as Soluble Pattern Recognition Receptors for Pathogenic Viruses
- PMID: 34063241
- PMCID: PMC8147407
- DOI: 10.3390/v13050824
Complement Proteins as Soluble Pattern Recognition Receptors for Pathogenic Viruses
Abstract
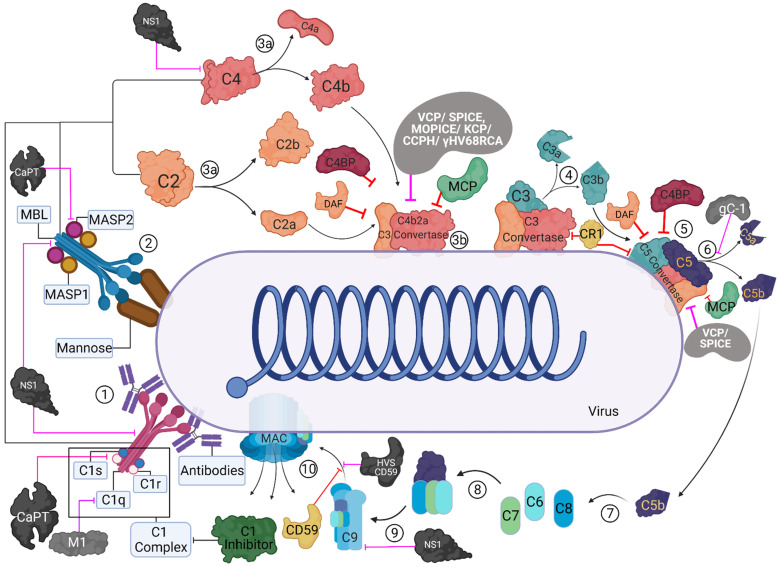
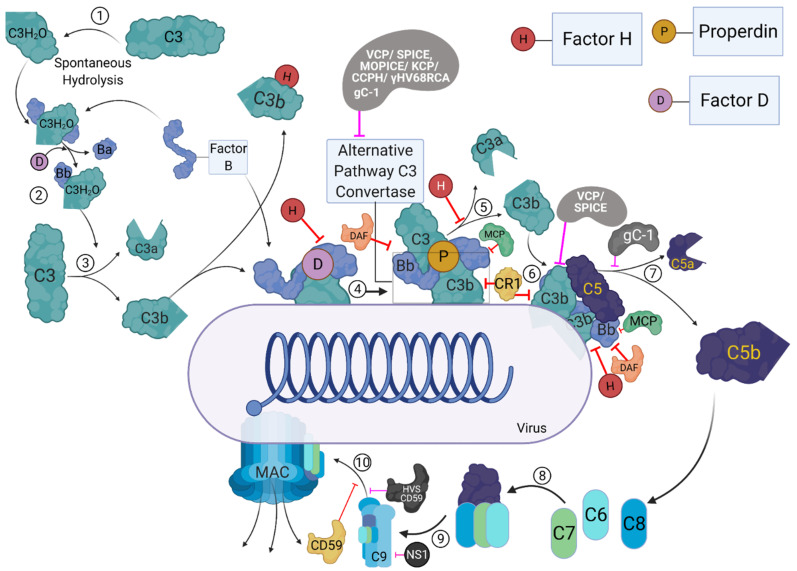
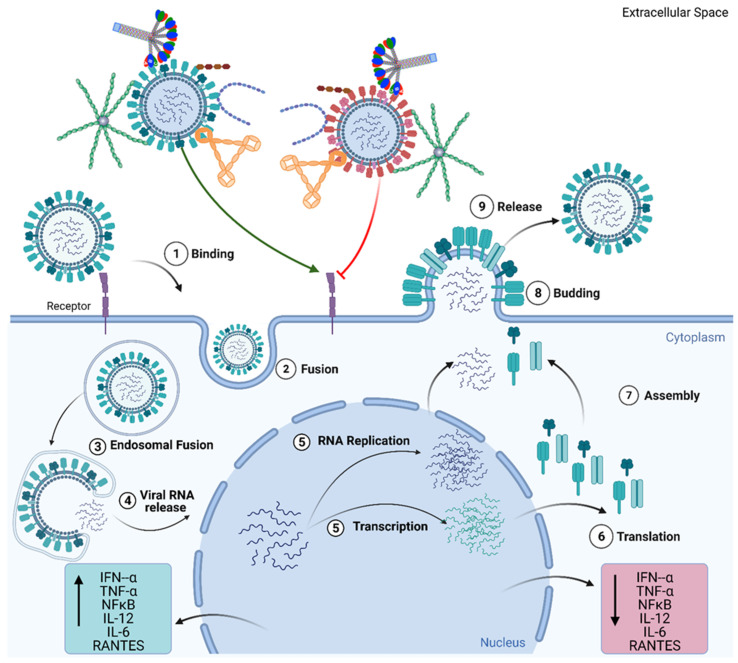
The complement system represents a crucial part of innate immunity. It contains a diverse range of soluble activators, membrane-bound receptors, and regulators. Its principal function is to eliminate pathogens via activation of three distinct pathways: classical, alternative, and lectin. In the case of viruses, the complement activation results in effector functions such as virion opsonisation by complement components, phagocytosis induction, virolysis by the membrane attack complex, and promotion of immune responses through anaphylatoxins and chemotactic factors. Recent studies have shown that the addition of individual complement components can neutralise viruses without requiring the activation of the complement cascade. While the complement-mediated effector functions can neutralise a diverse range of viruses, numerous viruses have evolved mechanisms to subvert complement recognition/activation by encoding several proteins that inhibit the complement system, contributing to viral survival and pathogenesis. This review focuses on these complement-dependent and -independent interactions of complement components (especially C1q, C4b-binding protein, properdin, factor H, Mannose-binding lectin, and Ficolins) with several viruses and their consequences.
Keywords: DNA viruses; RNA viruses; complement evasion; complement system; cytokine storm; innate immunity; retroviruses.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
