

Molecular Targets and Biological Functions of cAMP Signaling in Arabidopsis
- PMID: 34063698
- PMCID: PMC8147800
- DOI: 10.3390/biom11050688
Molecular Targets and Biological Functions of cAMP Signaling in Arabidopsis
Abstract
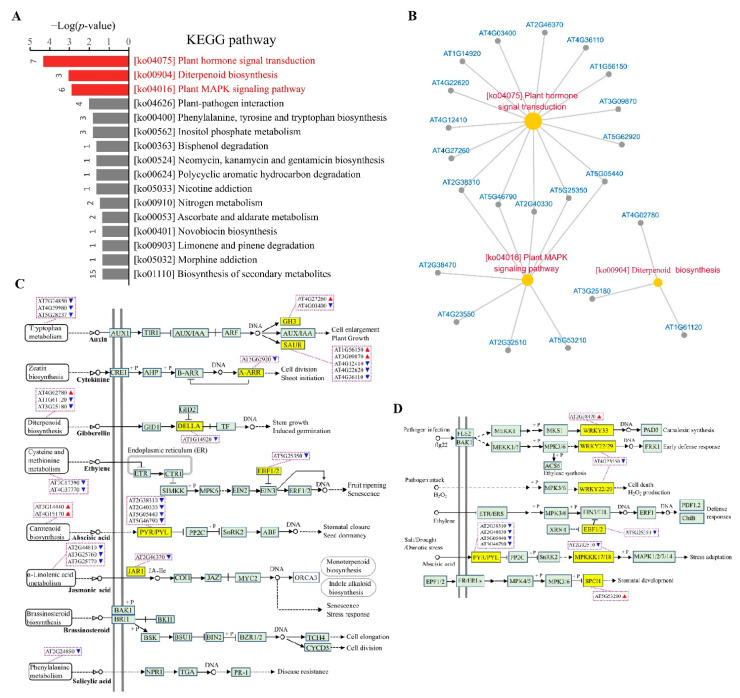
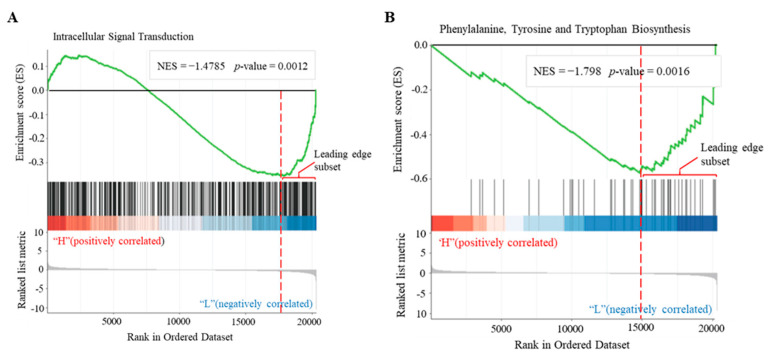
Cyclic AMP (cAMP) is a pivotal signaling molecule existing in almost all living organisms. However, the mechanism of cAMP signaling in plants remains very poorly understood. Here, we employ the engineered activity of soluble adenylate cyclase to induce cellular cAMP elevation in Arabidopsis thaliana plants and identify 427 cAMP-responsive genes (CRGs) through RNA-seq analysis. Induction of cellular cAMP elevation inhibits seed germination, disturbs phytohormone contents, promotes leaf senescence, impairs ethylene response, and compromises salt stress tolerance and pathogen resistance. A set of 62 transcription factors are among the CRGs, supporting a prominent role of cAMP in transcriptional regulation. The CRGs are significantly overrepresented in the pathways of plant hormone signal transduction, MAPK signaling, and diterpenoid biosynthesis, but they are also implicated in lipid, sugar, K+, nitrate signaling, and beyond. Our results provide a basic framework of cAMP signaling for the community to explore. The regulatory roles of cAMP signaling in plant plasticity are discussed.
Keywords: Arabidopsis thaliana; adenylate cyclase; cyclic AMP; function; mechanism; regulation; signaling.
Conflict of interest statement
The authors declare no conflict of interest.
Figures






Similar articles
-
The Arabidopsis DELAY OF GERMINATION 1 gene affects ABSCISIC ACID INSENSITIVE 5 (ABI5) expression and genetically interacts with ABI3 during Arabidopsis seed development.Plant J. 2016 Feb;85(4):451-65. doi: 10.1111/tpj.13118. Epub 2016 Feb 5. Plant J. 2016. PMID: 26729600
-
RSM1, an Arabidopsis MYB protein, interacts with HY5/HYH to modulate seed germination and seedling development in response to abscisic acid and salinity.PLoS Genet. 2018 Dec 19;14(12):e1007839. doi: 10.1371/journal.pgen.1007839. eCollection 2018 Dec. PLoS Genet. 2018. PMID: 30566447 Free PMC article.
-
Three Arabidopsis SnRK2 protein kinases, SRK2D/SnRK2.2, SRK2E/SnRK2.6/OST1 and SRK2I/SnRK2.3, involved in ABA signaling are essential for the control of seed development and dormancy.Plant Cell Physiol. 2009 Jul;50(7):1345-63. doi: 10.1093/pcp/pcp083. Epub 2009 Jun 18. Plant Cell Physiol. 2009. PMID: 19541597
-
Identification of two protein kinases required for abscisic acid regulation of seed germination, root growth, and gene expression in Arabidopsis.Plant Cell. 2007 Feb;19(2):485-94. doi: 10.1105/tpc.106.048538. Epub 2007 Feb 16. Plant Cell. 2007. PMID: 17307925 Free PMC article.
-
Endogenous cAMP elevation in Brassica napus causes changes in phytohormone levels.Plant Signal Behav. 2024 Dec 31;19(1):2310963. doi: 10.1080/15592324.2024.2310963. Epub 2024 Feb 5. Plant Signal Behav. 2024. PMID: 38314783 Free PMC article.
Cited by
-
GhCNGC13 and 32 Act as Critical Links between Growth and Immunity in Cotton.Int J Mol Sci. 2023 Dec 19;25(1):1. doi: 10.3390/ijms25010001. Int J Mol Sci. 2023. PMID: 38203172 Free PMC article.
-
The Identification of Key Genes and Biological Pathways in Heart Failure by Integrated Bioinformatics Analysis.Comput Math Methods Med. 2021 Nov 26;2021:3859338. doi: 10.1155/2021/3859338. eCollection 2021. Comput Math Methods Med. 2021. PMID: 34868339 Free PMC article.
-
Plant adenylate cyclases have come full circle.Nat Plants. 2023 Sep;9(9):1389-1397. doi: 10.1038/s41477-023-01486-x. Epub 2023 Sep 14. Nat Plants. 2023. PMID: 37709954 Review.
-
Functional Crypto-Adenylate Cyclases Operate in Complex Plant Proteins.Front Plant Sci. 2021 Aug 12;12:711749. doi: 10.3389/fpls.2021.711749. eCollection 2021. Front Plant Sci. 2021. PMID: 34456950 Free PMC article.
-
The cAMP-dependent phosphorylation footprint in response to heat stress.Plant Cell Rep. 2024 May 7;43(6):137. doi: 10.1007/s00299-024-03213-y. Plant Cell Rep. 2024. PMID: 38713285 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
