

α-Helices in the Type III Secretion Effectors: A Prevalent Feature with Versatile Roles
- PMID: 34063760
- PMCID: PMC8196651
- DOI: 10.3390/ijms22115412
α-Helices in the Type III Secretion Effectors: A Prevalent Feature with Versatile Roles
Abstract
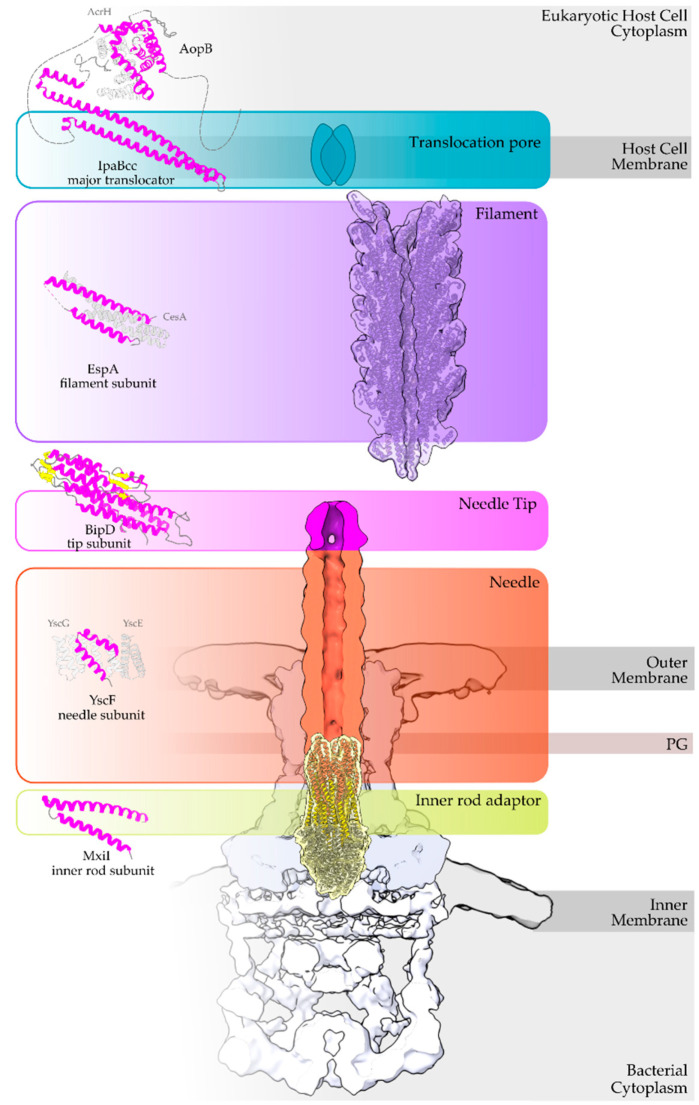
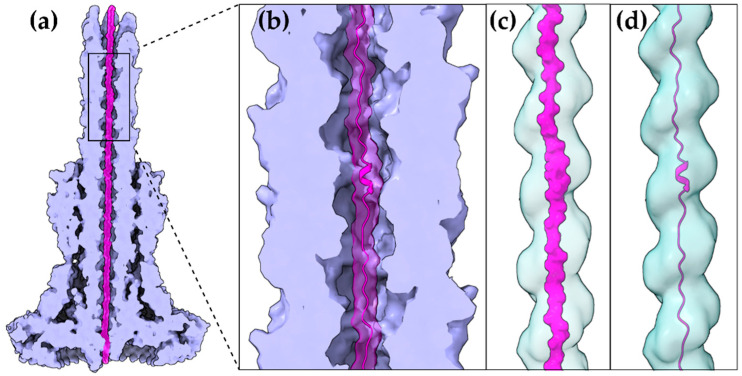
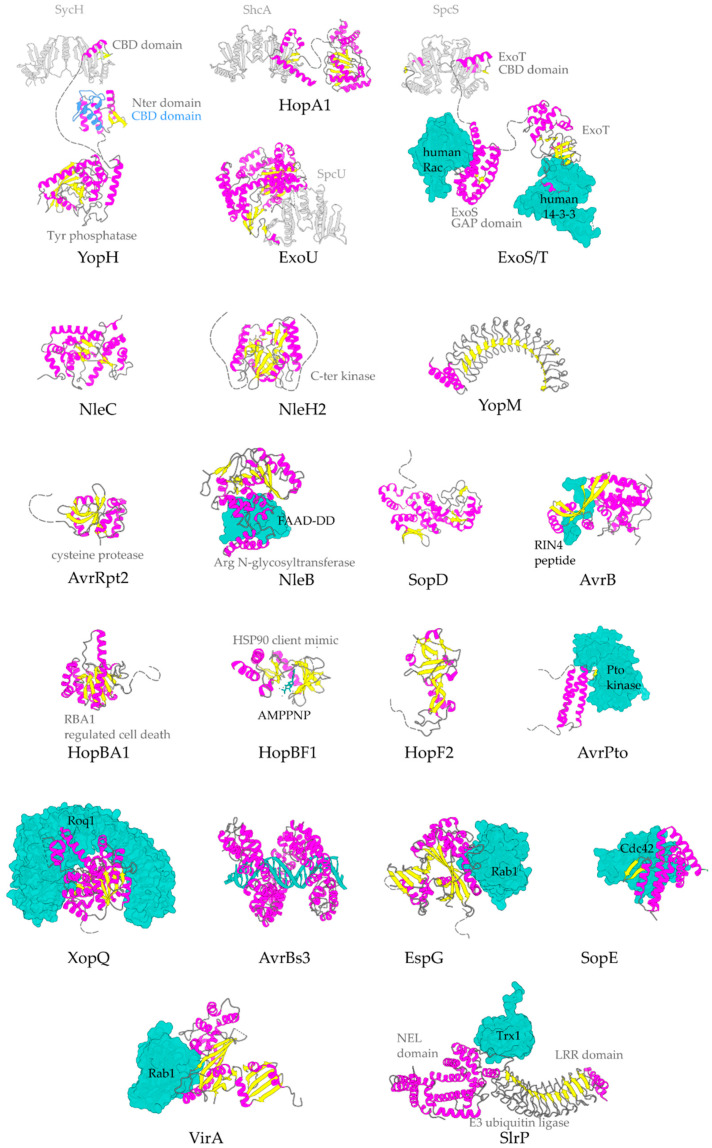
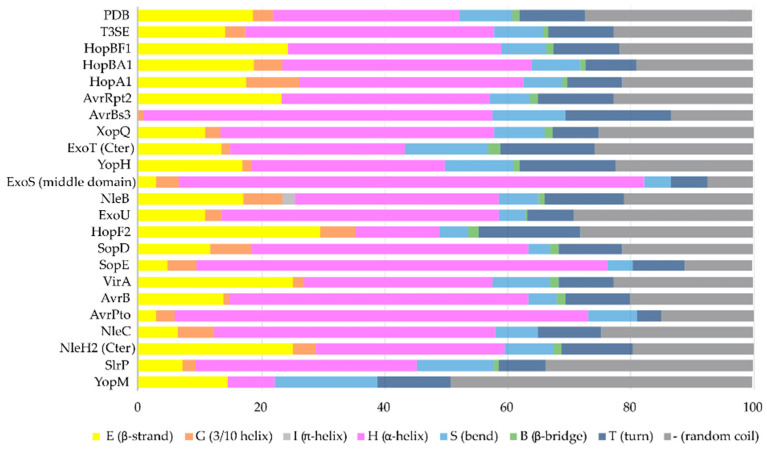
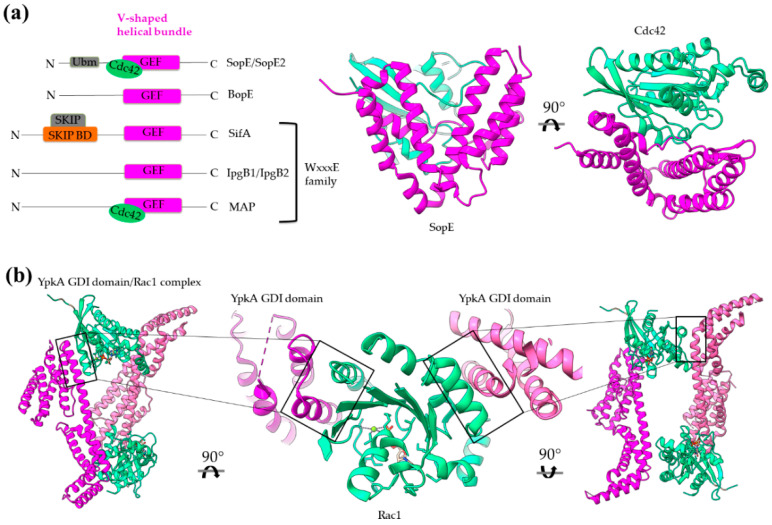
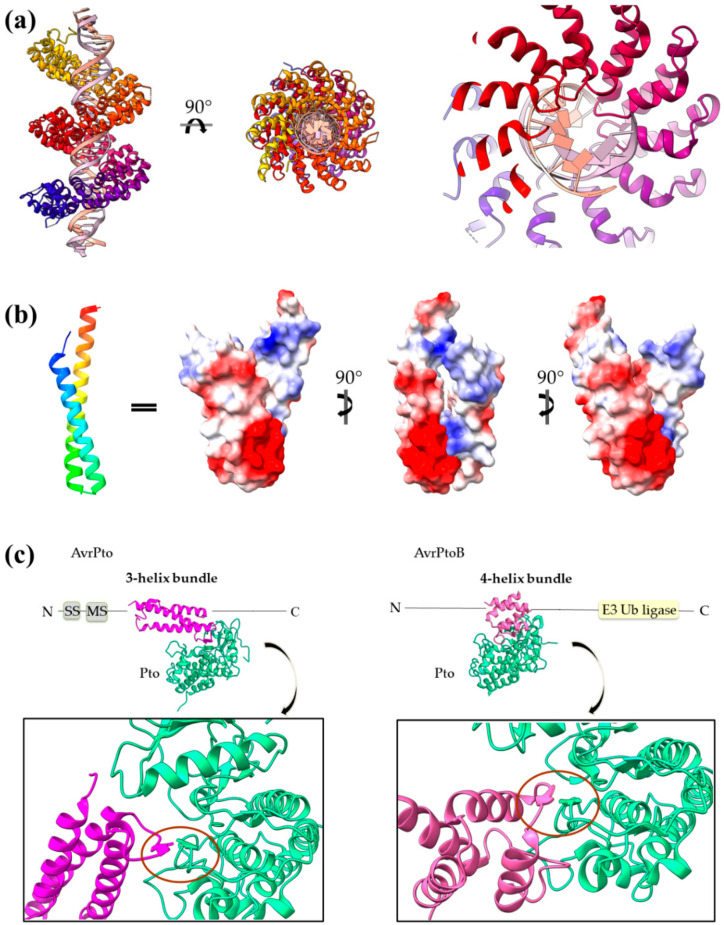
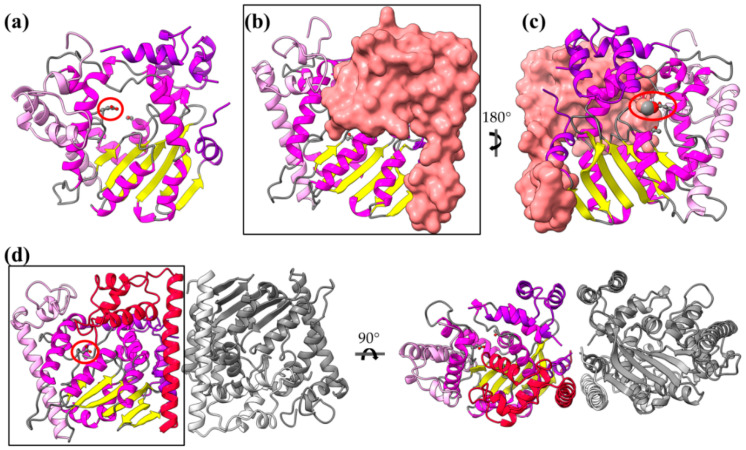
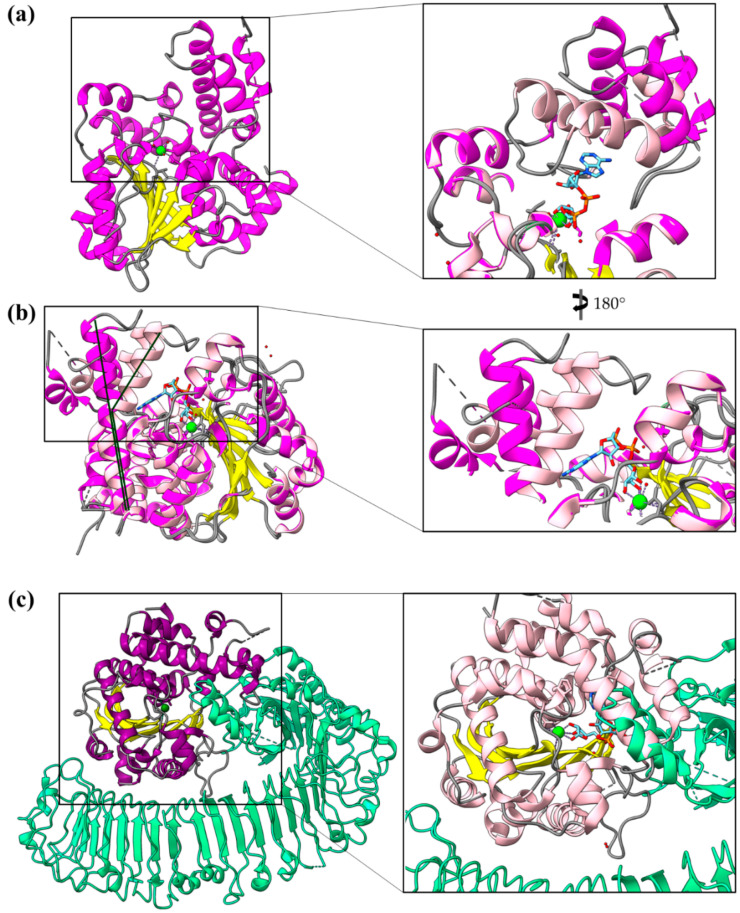
Type III Secretion Systems (T3SSs) are multicomponent nanomachines located at the cell envelope of Gram-negative bacteria. Their main function is to transport bacterial proteins either extracellularly or directly into the eukaryotic host cell cytoplasm. Type III Secretion effectors (T3SEs), latest to be secreted T3S substrates, are destined to act at the eukaryotic host cell cytoplasm and occasionally at the nucleus, hijacking cellular processes through mimicking eukaryotic proteins. A broad range of functions is attributed to T3SEs, ranging from the manipulation of the host cell's metabolism for the benefit of the bacterium to bypassing the host's defense mechanisms. To perform this broad range of manipulations, T3SEs have evolved numerous novel folds that are compatible with some basic requirements: they should be able to easily unfold, pass through the narrow T3SS channel, and refold to an active form when on the other side. In this review, the various folds of T3SEs are presented with the emphasis placed on the functional and structural importance of α-helices and helical domains.
Keywords: 4-α-helix bundle; Leucine-Rich Repeat (LRR); Novel E3 Ligase (NEL); Transcription Activator-Like Effector (TALE); Type III Secretion System (T3SS); Type III Secretion effector (T3SE); coiled coil; dictionary of secondary structure in proteins (DSSP).
Conflict of interest statement
The authors declare no conflict of interest.
Figures
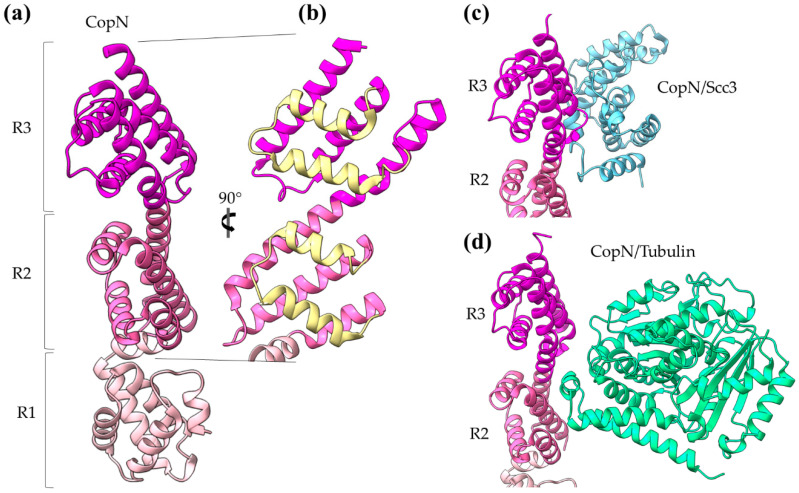
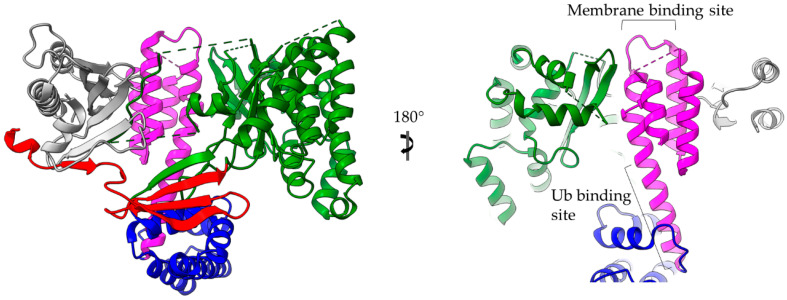
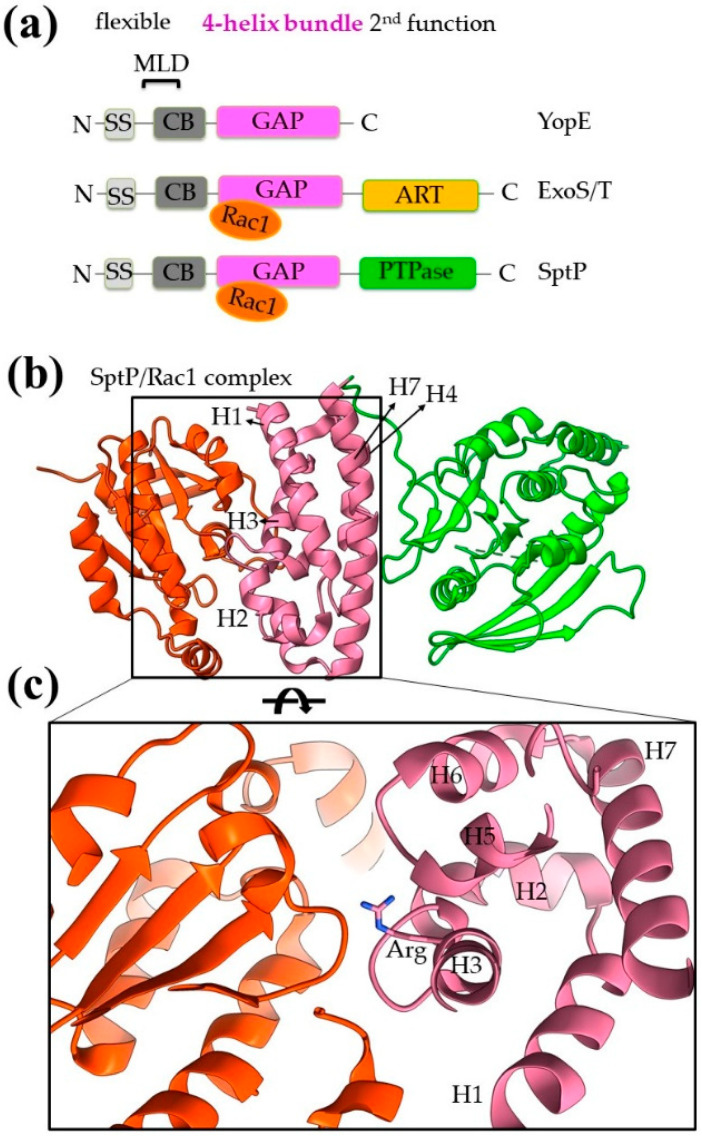
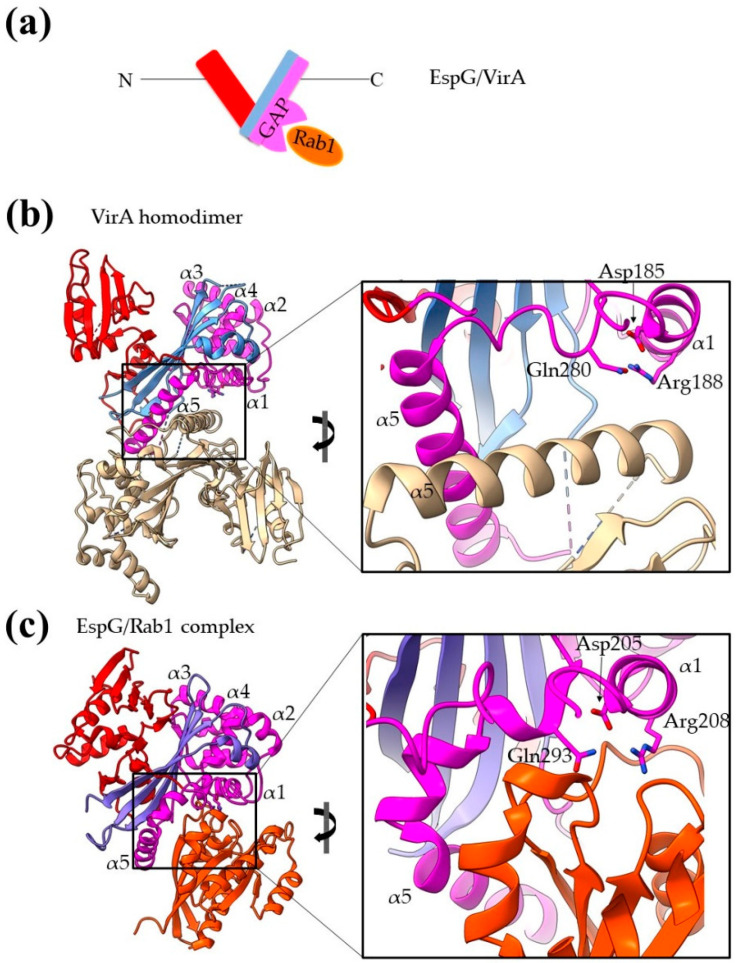
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
