

The AraC-Type Transcriptional Regulator GliR (PA3027) Activates Genes of Glycerolipid Metabolism in Pseudomonas aeruginosa
- PMID: 34064685
- PMCID: PMC8151288
- DOI: 10.3390/ijms22105066
The AraC-Type Transcriptional Regulator GliR (PA3027) Activates Genes of Glycerolipid Metabolism in Pseudomonas aeruginosa
Abstract
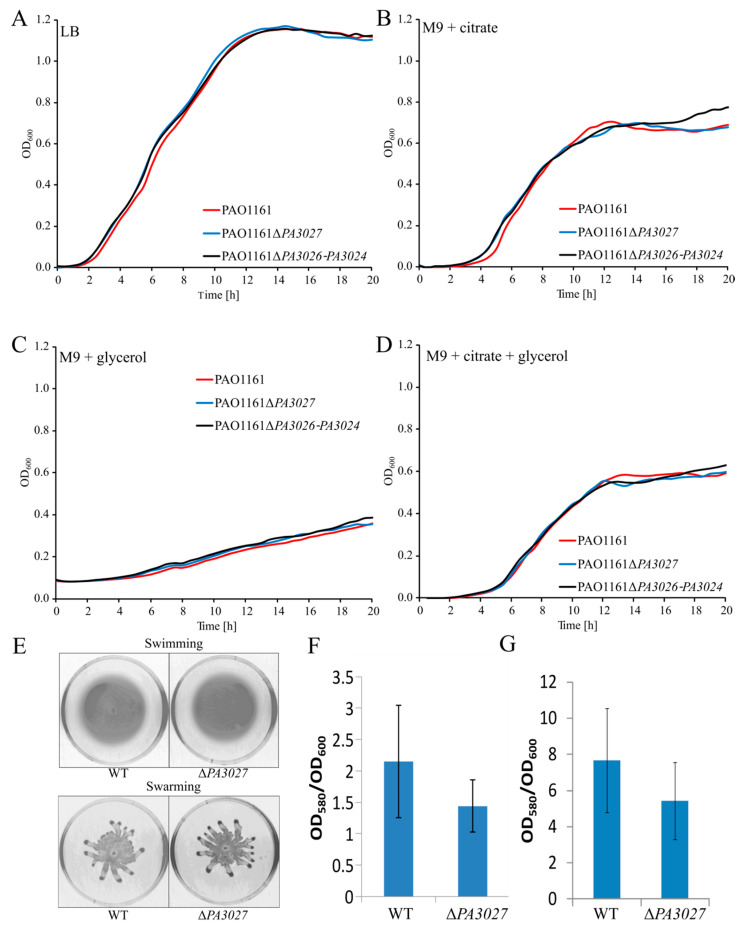
Pseudomonas aeruginosa encodes a large set of transcriptional regulators (TRs) that modulate and manage cellular metabolism to survive in variable environmental conditions including that of the human body. The AraC family regulators are an abundant group of TRs in bacteria, mostly acting as gene expression activators, controlling diverse cellular functions (e.g., carbon metabolism, stress response, and virulence). The PA3027 protein from P. aeruginosa has been classified in silico as a putative AraC-type TR. Transcriptional profiling of P. aeruginosa PAO1161 overexpressing PA3027 revealed a spectacular increase in the mRNA levels of PA3026-PA3024 (divergent to PA3027), PA3464, and PA3342 genes encoding proteins potentially involved in glycerolipid metabolism. Concomitantly, chromatin immunoprecipitation-sequencing (ChIP-seq) analysis revealed that at least 22 regions are bound by PA3027 in the PAO1161 genome. These encompass promoter regions of PA3026, PA3464, and PA3342, showing the major increase in expression in response to PA3027 excess. In Vitro DNA binding assay confirmed interactions of PA3027 with these regions. Furthermore, promoter-reporter assays in a heterologous host showed the PA3027-dependent activation of the promoter of the PA3026-PA3024 operon. Two motifs representing the preferred binding sites for PA3027, one localized upstream and one overlapping with the -35 promoter sequence, were identified in PA3026p and our data indicate that both motifs are required for full activation of this promoter by PA3027. Overall, the presented data show that PA3027 acts as a transcriptional regulator in P. aeruginosa, activating genes likely engaged in glycerolipid metabolism. The GliR name, from a glycerolipid metabolism regulator, is proposed for PA3027 of P. aeruginosa.
Keywords: AraC family; PA3027; Pseudomonas aeruginosa; glycerolipid metabolism; regulon; transcriptional regulator.
Conflict of interest statement
The authors declare no conflict of interest.
Figures







References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
