

SUMO-Targeted Ubiquitin Ligases and Their Functions in Maintaining Genome Stability
- PMID: 34065507
- PMCID: PMC8161396
- DOI: 10.3390/ijms22105391
SUMO-Targeted Ubiquitin Ligases and Their Functions in Maintaining Genome Stability
Abstract
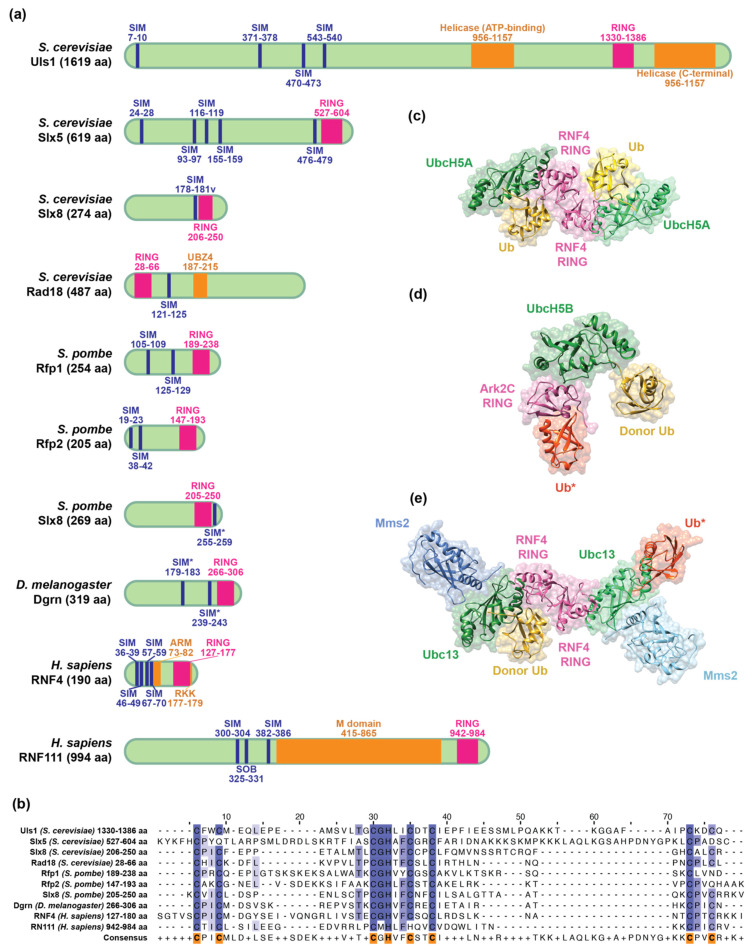
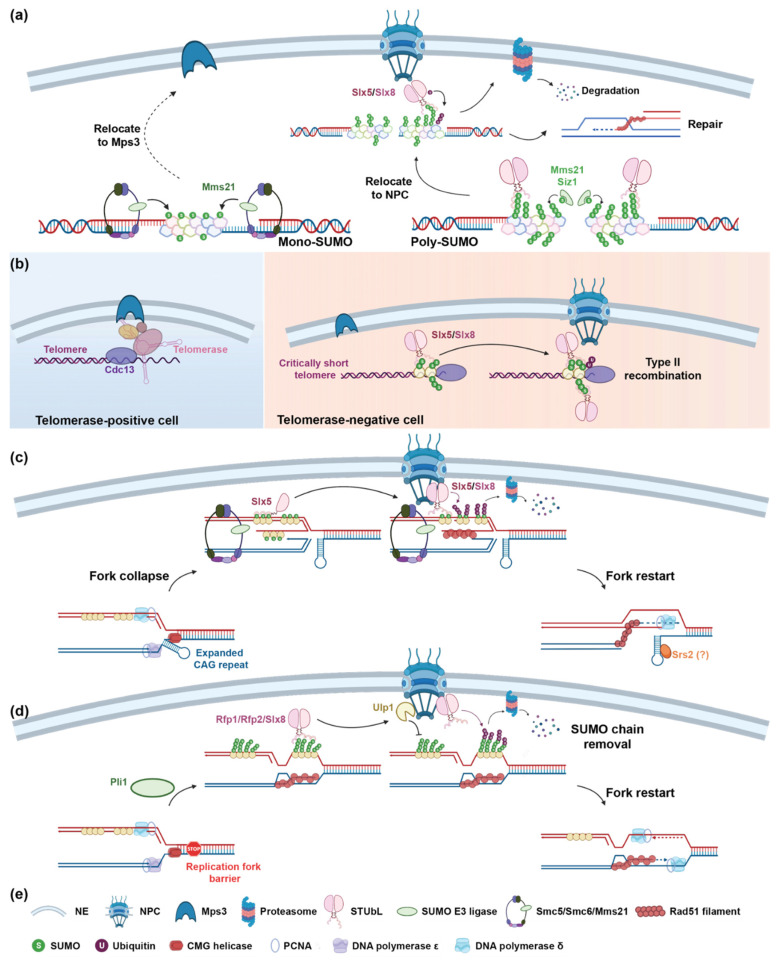
Small ubiquitin-like modifier (SUMO)-targeted E3 ubiquitin ligases (STUbLs) are specialized enzymes that recognize SUMOylated proteins and attach ubiquitin to them. They therefore connect the cellular SUMOylation and ubiquitination circuits. STUbLs participate in diverse molecular processes that span cell cycle regulated events, including DNA repair, replication, mitosis, and transcription. They operate during unperturbed conditions and in response to challenges, such as genotoxic stress. These E3 ubiquitin ligases modify their target substrates by catalyzing ubiquitin chains that form different linkages, resulting in proteolytic or non-proteolytic outcomes. Often, STUbLs function in compartmentalized environments, such as the nuclear envelope or kinetochore, and actively aid in nuclear relocalization of damaged DNA and stalled replication forks to promote DNA repair or fork restart. Furthermore, STUbLs reside in the same vicinity as SUMO proteases and deubiquitinases (DUBs), providing spatiotemporal control of their targets. In this review, we focus on the molecular mechanisms by which STUbLs help to maintain genome stability across different species.
Keywords: RNF111; RNF4; STUbL; SUMO; Slx5/Slx8; genome stability; ubiquitin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Preserving genome integrity: The vital role of SUMO-targeted ubiquitin ligases.Cell Insight. 2023 Oct 23;2(6):100128. doi: 10.1016/j.cellin.2023.100128. eCollection 2023 Dec. Cell Insight. 2023. PMID: 38047137 Free PMC article. Review.
-
SUMO-targeted ubiquitin ligases in genome stability.EMBO J. 2007 Sep 19;26(18):4089-101. doi: 10.1038/sj.emboj.7601838. Epub 2007 Aug 30. EMBO J. 2007. PMID: 17762865 Free PMC article.
-
RNF111/Arkadia is a SUMO-targeted ubiquitin ligase that facilitates the DNA damage response.J Cell Biol. 2013 Jun 10;201(6):797-807. doi: 10.1083/jcb.201212075. J Cell Biol. 2013. PMID: 23751493 Free PMC article.
-
Methods to analyze STUbL activity.Methods Enzymol. 2019;618:257-280. doi: 10.1016/bs.mie.2018.11.005. Epub 2019 Jan 3. Methods Enzymol. 2019. PMID: 30850055
-
A SIM-ultaneous role for SUMO and ubiquitin.Trends Biochem Sci. 2008 May;33(5):201-8. doi: 10.1016/j.tibs.2008.02.001. Epub 2008 Apr 9. Trends Biochem Sci. 2008. PMID: 18403209 Review.
Cited by
-
RNF4 and USP7 cooperate in ubiquitin-regulated steps of DNA replication.Open Biol. 2023 Aug;13(8):230068. doi: 10.1098/rsob.230068. Epub 2023 Aug 23. Open Biol. 2023. PMID: 37607592 Free PMC article.
-
Sequential post-translational modifications regulate damaged DNA-binding protein DDB2 function.J Biochem. 2024 Sep 30;176(4):325-338. doi: 10.1093/jb/mvae056. J Biochem. 2024. PMID: 39077792 Free PMC article.
-
Preserving genome integrity: The vital role of SUMO-targeted ubiquitin ligases.Cell Insight. 2023 Oct 23;2(6):100128. doi: 10.1016/j.cellin.2023.100128. eCollection 2023 Dec. Cell Insight. 2023. PMID: 38047137 Free PMC article. Review.
-
The Intra- and Extra-Telomeric Role of TRF2 in the DNA Damage Response.Int J Mol Sci. 2021 Sep 14;22(18):9900. doi: 10.3390/ijms22189900. Int J Mol Sci. 2021. PMID: 34576063 Free PMC article. Review.
-
Regulating the p53 Tumor Suppressor Network at PML Biomolecular Condensates.Cancers (Basel). 2022 Sep 20;14(19):4549. doi: 10.3390/cancers14194549. Cancers (Basel). 2022. PMID: 36230470 Free PMC article. Review.
References
-
- Kotsantis P., Petermann E., Boulton S.J. Mechanisms of Oncogene-Induced Replication Stress: Jigsaw Falling into Place. Cancer Discov. 2018;8:537–555. doi: 10.1158/2159-8290.CD-17-1461. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
