

PERK Is Critical for Alphavirus Nonstructural Protein Translation
- PMID: 34065980
- PMCID: PMC8151226
- DOI: 10.3390/v13050892
PERK Is Critical for Alphavirus Nonstructural Protein Translation
Retraction in
-
RETRACTED: Dahal et al. PERK Is Critical for Alphavirus Nonstructural Protein Translation. Viruses 2021, 13, 892.Viruses. 2025 Feb 26;17(3):318. doi: 10.3390/v17030318. Viruses. 2025. PMID: 40143380 Free PMC article.
Abstract
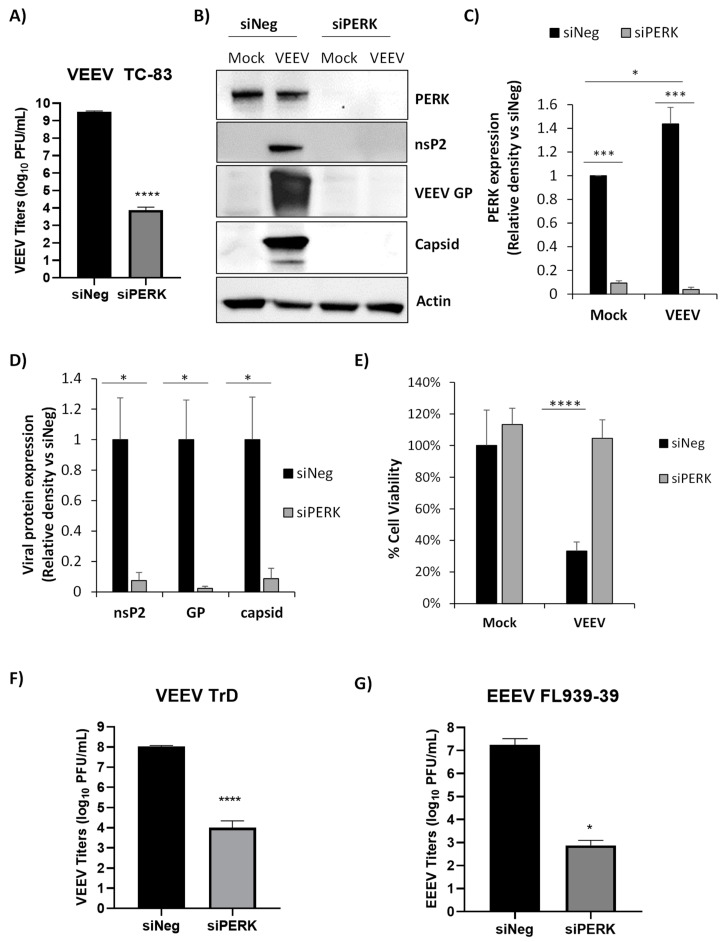
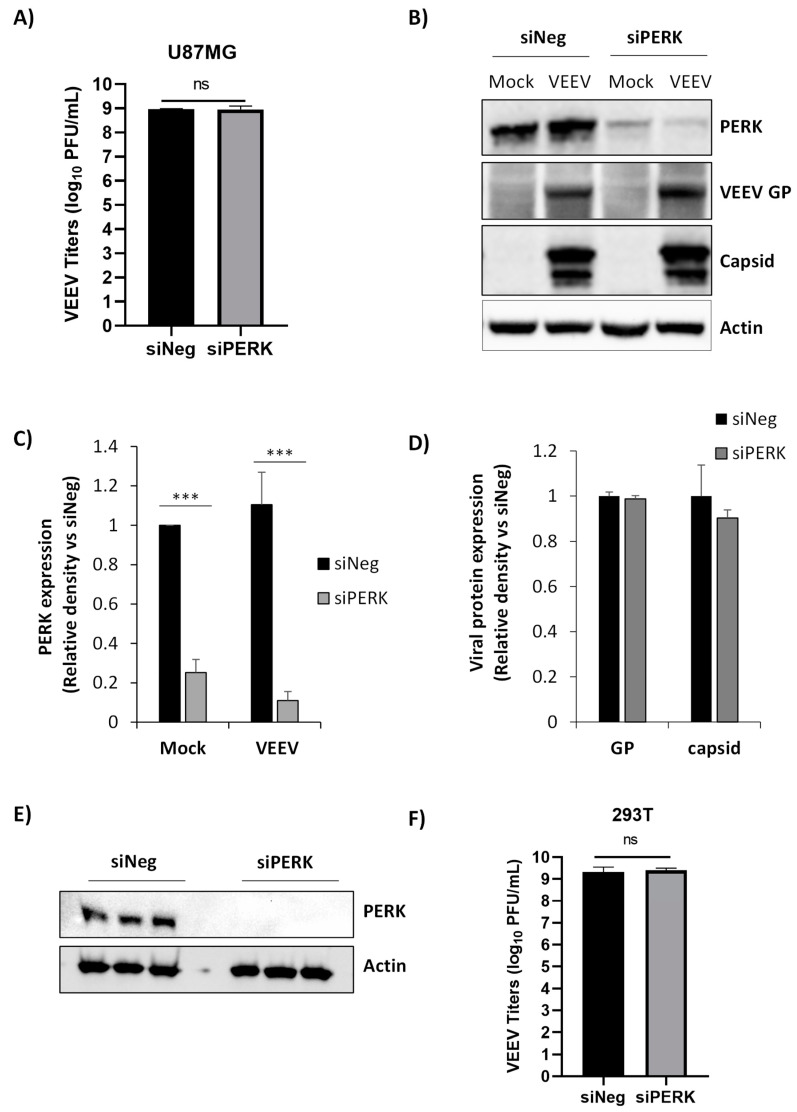
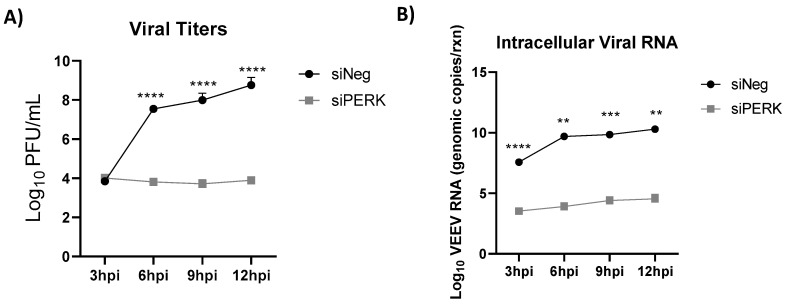
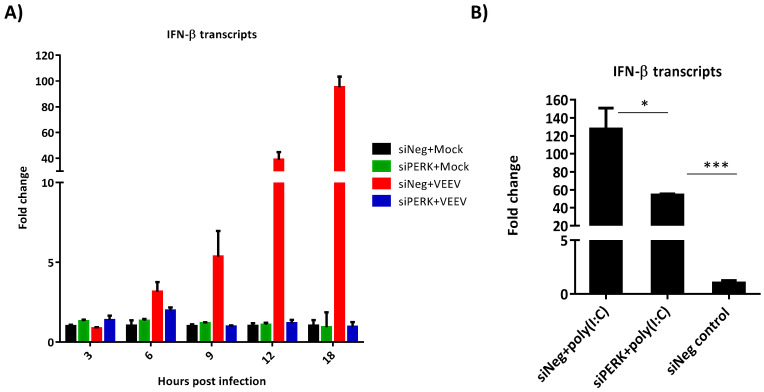
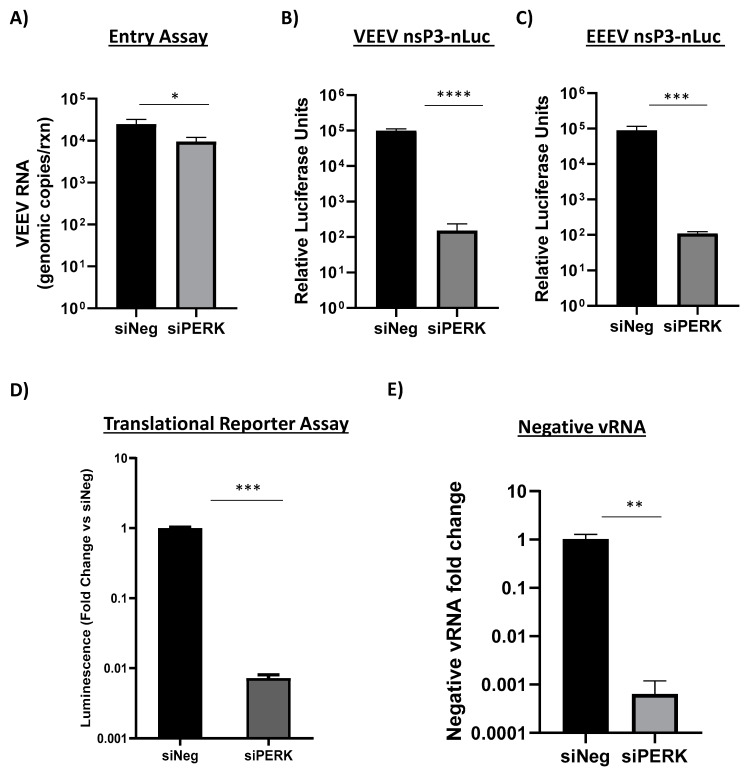
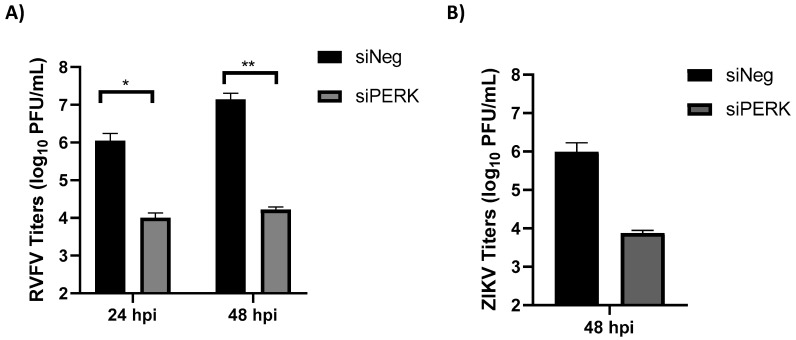
Venezuelan equine encephalitis virus (VEEV) is an alphavirus that causes encephalitis. Previous work indicated that VEEV infection induced early growth response 1 (EGR1) expression, leading to cell death via the protein kinase R (PKR)-like endoplasmic reticulum kinase (PERK) arm of the unfolded protein response (UPR) pathway. Loss of PERK prevented EGR1 induction and decreased VEEV-induced death. The results presented within show that loss of PERK in human primary astrocytes dramatically reduced VEEV and eastern equine encephalitis virus (EEEV) infectious titers by 4-5 log10. Loss of PERK also suppressed VEEV replication in primary human pericytes and human umbilical vein endothelial cells, but it had no impact on VEEV replication in transformed U87MG and 293T cells. A significant reduction in VEEV RNA levels was observed as early as 3 h post-infection, but viral entry assays indicated that the loss of PERK minimally impacted VEEV entry. In contrast, the loss of PERK resulted in a dramatic reduction in viral nonstructural protein translation and negative-strand viral RNA production. The loss of PERK also reduced the production of Rift Valley fever virus and Zika virus infectious titers. These data indicate that PERK is an essential factor for the translation of alphavirus nonstructural proteins and impacts multiple RNA viruses, making it an exciting target for antiviral development.
Keywords: PERK; Venezuelan equine encephalitis virus (VEEV); alphavirus; eastern equine encephalitis virus (EEEV); translation.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures
Similar articles
-
EGR1 upregulation following Venezuelan equine encephalitis virus infection is regulated by ERK and PERK pathways contributing to cell death.Virology. 2020 Jan 2;539:121-128. doi: 10.1016/j.virol.2019.10.016. Epub 2019 Oct 31. Virology. 2020. PMID: 31733451 Free PMC article.
-
Venezuelan Equine Encephalitis Virus Induces Apoptosis through the Unfolded Protein Response Activation of EGR1.J Virol. 2016 Jan 20;90(7):3558-72. doi: 10.1128/JVI.02827-15. J Virol. 2016. PMID: 26792742 Free PMC article.
-
Host translation shutoff mediated by non-structural protein 2 is a critical factor in the antiviral state resistance of Venezuelan equine encephalitis virus.Virology. 2016 Sep;496:147-165. doi: 10.1016/j.virol.2016.06.005. Epub 2016 Jun 15. Virology. 2016. PMID: 27318152 Free PMC article.
-
Venezuelan Equine Encephalitis Virus Capsid-The Clever Caper.Viruses. 2017 Sep 29;9(10):279. doi: 10.3390/v9100279. Viruses. 2017. PMID: 28961161 Free PMC article. Review.
-
Role of Angiotensin II in Venezuelan Equine Encephalitis: Narrative Review.Rev Med Virol. 2025 May;35(3):e70040. doi: 10.1002/rmv.70040. Rev Med Virol. 2025. PMID: 40317791 Review.
Cited by
-
Interactions of Equine Viruses with the Host Kinase Machinery and Implications for One Health and Human Disease.Viruses. 2023 May 13;15(5):1163. doi: 10.3390/v15051163. Viruses. 2023. PMID: 37243249 Free PMC article. Review.
-
Cellular Stress Responses against Coronavirus Infection: A Means of the Innate Antiviral Defense.J Microbiol Biotechnol. 2024 Jan 28;34(1):1-9. doi: 10.4014/jmb.2307.07038. Epub 2023 Sep 8. J Microbiol Biotechnol. 2024. PMID: 37674398 Free PMC article. Review.
-
The role of endoplasmic reticulum stress in astrocytes.Glia. 2022 Jan;70(1):5-19. doi: 10.1002/glia.24082. Epub 2021 Aug 31. Glia. 2022. PMID: 34462963 Free PMC article. Review.
-
Endoplasmic reticulum stress in diseases.MedComm (2020). 2024 Aug 26;5(9):e701. doi: 10.1002/mco2.701. eCollection 2024 Sep. MedComm (2020). 2024. PMID: 39188936 Free PMC article. Review.
-
Emerging mechanisms of the unfolded protein response in therapeutic resistance: from chemotherapy to Immunotherapy.Cell Commun Signal. 2024 Jan 31;22(1):89. doi: 10.1186/s12964-023-01438-0. Cell Commun Signal. 2024. PMID: 38297380 Free PMC article. Review.
References
-
- Gardner C.L., Burke C.W., Tesfay M.Z., Glass P.J., Klimstra W.B., Ryman K.D. Eastern and Venezuelan equine encephalitis viruses differ in their ability to infect dendritic cells and macrophages: Impact of altered cell tropism on pathogenesis. J. Virol. 2008;82:10634–10646. doi: 10.1128/JVI.01323-08. - DOI - PMC - PubMed
-
- Dahal B., Lin S.-C., Carey B.D., Jacobs J.L., Dinman J.D., van Hoek M.L., Adams A.A., Kehn-Hall K. EGR1 upregulation following Venezuelan equine encephalitis virus infection is regulated by ERK and PERK pathways contributing to cell death. Virology. 2019 doi: 10.1016/j.virol.2019.10.016. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
