

Structural Plasticity of LL-37 Indicates Elaborate Functional Adaptation Mechanisms to Bacterial Target Structures
- PMID: 34068993
- PMCID: PMC8156758
- DOI: 10.3390/ijms22105200
Structural Plasticity of LL-37 Indicates Elaborate Functional Adaptation Mechanisms to Bacterial Target Structures
Abstract
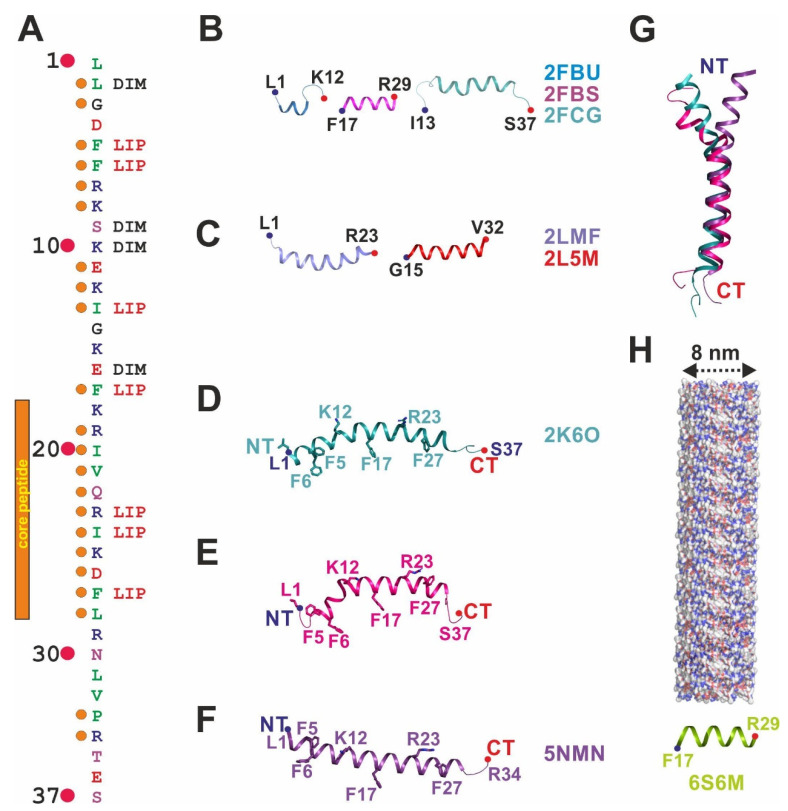
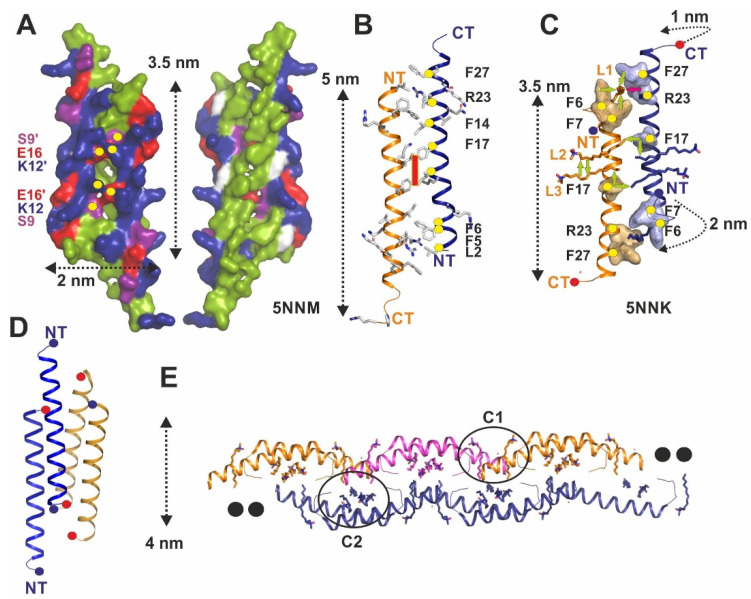
The human cathelicidin LL-37 is a multifunctional peptide of the human innate immune system. Among the various functions of LL-37, its antimicrobial activity is important in controlling the microorganisms of the human body. The target molecules of LL-37 in bacteria include membrane lipids, lipopolysaccharides (LPS), lipoteichoic acid (LTA), proteins, DNA and RNA. In this mini-review, we summarize the entity of LL-37 structural data determined over the last 15 years and specifically discuss features implicated in the interactions with lipid-like molecules. For this purpose, we discuss partial and full-length structures of LL-37 determined in the presence of membrane-mimicking detergents. This constantly growing structural database is now composed of monomers, dimers, tetramers, and fiber-like structures. The diversity of these structures underlines an unexpected plasticity and highlights the conformational and oligomeric adaptability of LL-37 necessary to target different molecular scaffolds. The recent co-crystal structures of LL-37 in complex with detergents are particularly useful to understand how these molecules mimic lipids and LPS to induce oligomerization and fibrillation. Defined detergent binding sites provide deep insights into a new class of peptide scaffolds, widening our view on the fascinating world of the LL-37 structural factotum. Together, the new structures in their evolutionary context allow for the assignment of functionally conserved residues in oligomerization and target interactions. Conserved phenylalanine and arginine residues primarily mediate those interactions with lipids and LPS. The interactions with macromolecules such as proteins or DNA remain largely unexplored and open a field for future studies aimed at structures of LL-37 complexes. These complexes will then allow for the structure-based rational design of LL-37-derived peptides with improved antibiotic properties.
Keywords: LL-37; antimicrobial peptides; cathelicidin; structural flexibility.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Koo H.B., Seo J. Antimicrobial peptides under clinical investigation. Pept. Sci. 2019;111:e24122. doi: 10.1002/pep2.24122. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
