

A MADS-Box Gene CiMADS43 Is Involved in Citrus Flowering and Leaf Development through Interaction with CiAGL9
- PMID: 34069068
- PMCID: PMC8156179
- DOI: 10.3390/ijms22105205
A MADS-Box Gene CiMADS43 Is Involved in Citrus Flowering and Leaf Development through Interaction with CiAGL9
Abstract
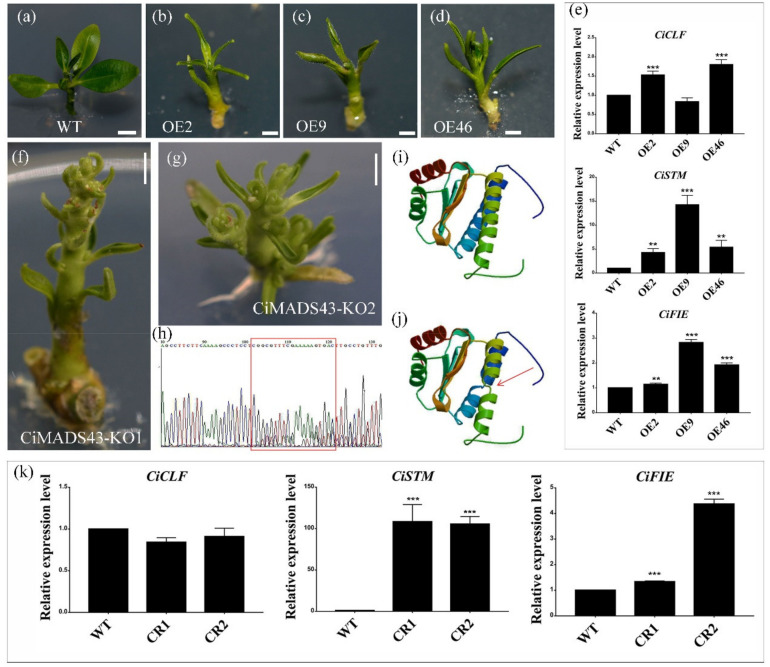
MADS-box genes are involved in various developmental processes including vegetative development, flower architecture, flowering, pollen formation, seed and fruit development. However, the function of most MADS-box genes and their regulation mechanism are still unclear in woody plants compared with model plants. In this study, a MADS-box gene (CiMADS43) was identified in citrus. Phylogenetic and sequence analysis showed that CiMADS43 is a GOA-like Bsister MADS-box gene. It was localized in the nucleus and as a transcriptional activator. Overexpression of CiMADS43 promoted early flowering and leaves curling in transgenic Arabidopsis. Besides, overexpression or knockout of CiMADS43 also showed leaf curl phenotype in citrus similar to that of CiMADS43 overexpressed in Arabidopsis. Protein-protein interaction found that a SEPALLATA (SEP)-like protein (CiAGL9) interacted with CiMADS43 protein. Interestingly, CiAGL9 also can bind to the CiMADS43 promoter and promote its transcription. Expression analysis also showed that these two genes were closely related to seasonal flowering and the development of the leaf in citrus. Our findings revealed the multifunctional roles of CiMADS43 in the vegetative and reproductive development of citrus. These results will facilitate our understanding of the evolution and molecular mechanisms of MADS-box genes in citrus.
Keywords: CiAGL9; CiMADS43; citrus; flowering; leaf.
Conflict of interest statement
The authors have no conflict of interest to declare.
Figures







Similar articles
-
Functional and evolutionary analysis of the AP1/SEP/AGL6 superclade of MADS-box genes in the basal eudicot Epimedium sagittatum.Ann Bot. 2014 Mar;113(4):653-68. doi: 10.1093/aob/mct301. Epub 2014 Feb 13. Ann Bot. 2014. PMID: 24532606 Free PMC article.
-
Ectopic expression of the petunia MADS box gene UNSHAVEN accelerates flowering and confers leaf-like characteristics to floral organs in a dominant-negative manner.Plant Cell. 2004 Jun;16(6):1490-505. doi: 10.1105/tpc.019679. Epub 2004 May 21. Plant Cell. 2004. PMID: 15155884 Free PMC article.
-
Overexpression of the MADS-box gene K-domain increases the yield potential of blueberry.Plant Sci. 2018 Nov;276:22-31. doi: 10.1016/j.plantsci.2018.07.018. Epub 2018 Jul 31. Plant Sci. 2018. PMID: 30348321
-
Make hay when the sun shines: the role of MADS-box genes in temperature-dependant seasonal flowering responses.Plant Sci. 2011 Mar;180(3):447-53. doi: 10.1016/j.plantsci.2010.12.001. Epub 2010 Dec 14. Plant Sci. 2011. PMID: 21421391 Review.
-
MADS-box genes and floral development: the dark side.J Exp Bot. 2012 Sep;63(15):5397-404. doi: 10.1093/jxb/ers233. Epub 2012 Aug 21. J Exp Bot. 2012. PMID: 22915743 Review.
Cited by
-
Genome-Wide Identification of MADS-Box Genes in Taraxacum kok-saghyz and Taraxacum mongolicum: Evolutionary Mechanisms, Conserved Functions and New Functions Related to Natural Rubber Yield Formation.Int J Mol Sci. 2023 Jul 1;24(13):10997. doi: 10.3390/ijms241310997. Int J Mol Sci. 2023. PMID: 37446175 Free PMC article.
-
Evolution and Function of MADS-Box Transcription Factors in Plants.Int J Mol Sci. 2024 Dec 11;25(24):13278. doi: 10.3390/ijms252413278. Int J Mol Sci. 2024. PMID: 39769043 Free PMC article. Review.
-
Transcriptome analysis of Harumi tangor fruits: Insights into interstock-mediated fruit quality.Front Plant Sci. 2022 Oct 13;13:995913. doi: 10.3389/fpls.2022.995913. eCollection 2022. Front Plant Sci. 2022. PMID: 36311145 Free PMC article.
-
Transcriptomic profiling and gene network analysis revealed regulatory mechanisms of bract development in Bougainvillea glabra.BMC Plant Biol. 2024 Jun 13;24(1):543. doi: 10.1186/s12870-024-05246-7. BMC Plant Biol. 2024. PMID: 38872082 Free PMC article.
-
Genome-wide identification of the AcMADS-box family and functional validation of AcMADS32 involved in carotenoid biosynthesis in Actinidia.Front Plant Sci. 2023 Jun 19;14:1159942. doi: 10.3389/fpls.2023.1159942. eCollection 2023. Front Plant Sci. 2023. PMID: 37404538 Free PMC article.
References
-
- Saha G., Park J.I., Jung H.J., Ahmed N.U., Kayum M.A., Chung M.Y., Hur Y., Cho Y.G., Watanabe M., Nou I.S. Genome-wide identification and characterization of MADS-box family genes related to organ development and stress resistance in Brassica rapa. BMC Genom. 2015;16:178. doi: 10.1186/s12864-015-1349-z. - DOI - PMC - PubMed
-
- Alvarez-Buylla E.R., Pelaz S., Liljegren S.J., Gold S.E., Burgeff C., Ditta G.S., de Pouplana L.R., Martinez-Castilla L., Yanofsky M.F. An ancestral MADS-box gene duplication occurred before the divergence of plants and animals. Proc. Natl. Acad. Sci. USA. 2000;97:5328–5333. doi: 10.1073/pnas.97.10.5328. - DOI - PMC - PubMed
-
- Parenicova L., de Folter S., Kieffer M., Horner D.S., Favalli C., Busscher J., Cook H.E., Ingram R.M., Kater M.M., Davies B., et al. Molecular and phylogenetic analyses of the complete MADS-box transcription factor family in Arabidopsis: New openings to the MADS world. Plant Cell. 2003;15:1538–1551. doi: 10.1105/tpc.011544. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
