

A New Mechanism in THRA Resistance: The First Disease-Associated Variant Leading to an Increased Inhibitory Function of THRA2
- PMID: 34069457
- PMCID: PMC8159125
- DOI: 10.3390/ijms22105338
A New Mechanism in THRA Resistance: The First Disease-Associated Variant Leading to an Increased Inhibitory Function of THRA2
Abstract
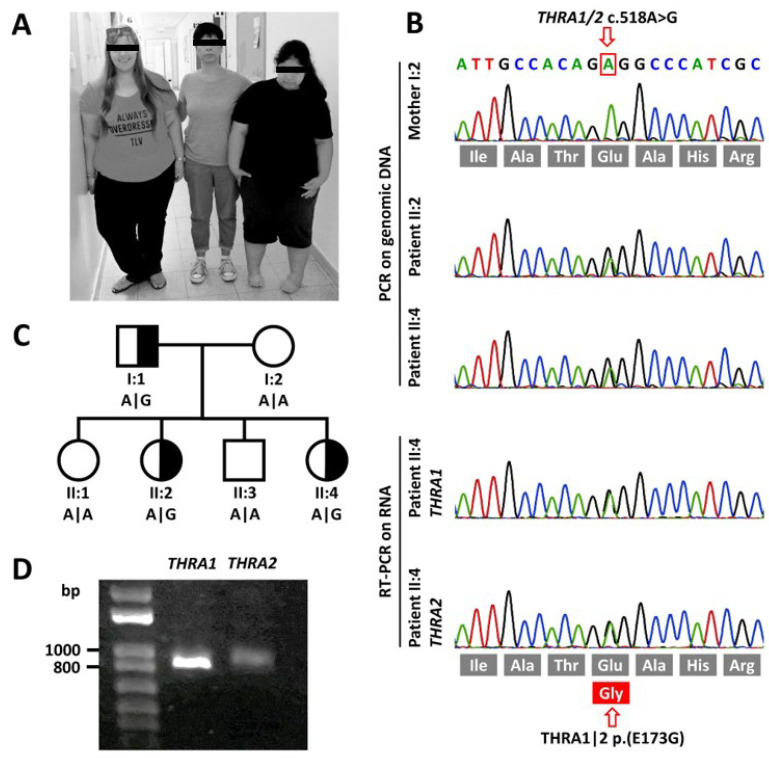
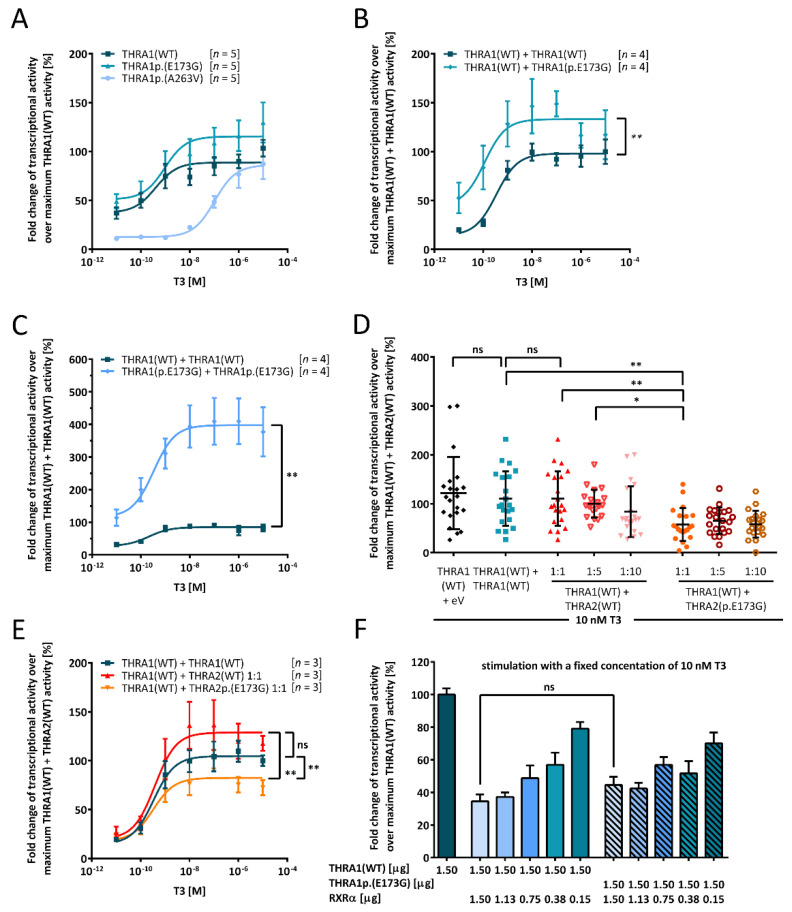
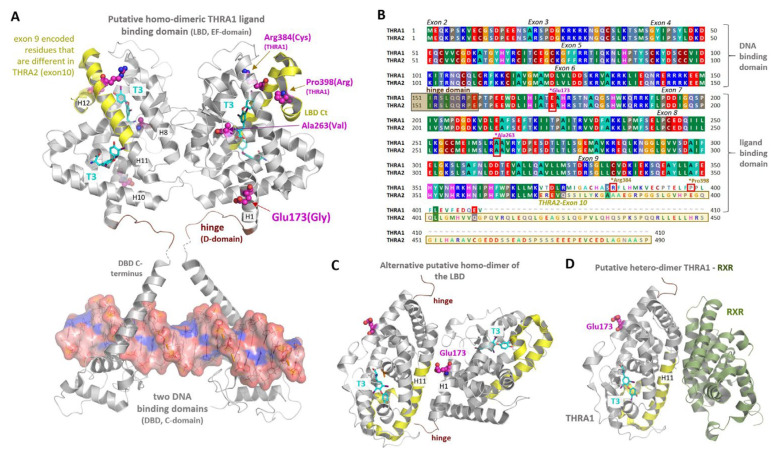
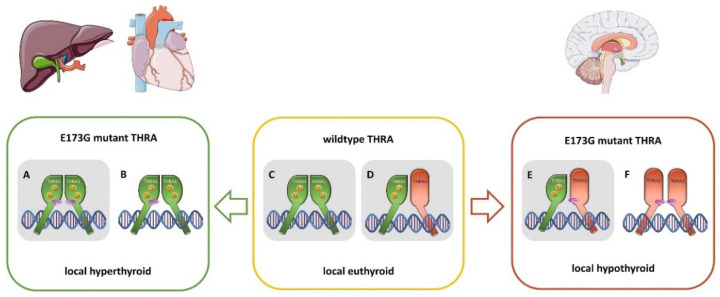
The nuclear thyroid hormone receptors (THRs) are key mediators of thyroid hormone function on the cellular level via modulation of gene expression. Two different genes encode THRs (THRA and THRB), and are pleiotropically involved in development, metabolism, and growth. The THRA1 and THRA2 isoforms, which result from alternative splicing of THRA, differ in their C-terminal ligand-binding domain (LBD). Most published disease-associated THRA variants are located in the LBD of THRA1 and impede triiodothyronine (T3) binding. This keeps the nuclear receptor in an inactive state and inhibits target gene expression. Here, we investigated a new dominant THRA variant (chr17:g.38,241,010A > G, GRCh37.13 | c.518A > G, NM_199334 | p.(E173G), NP_955366), which is located between the DNA- and ligand-binding domains and affects both splicing isoforms. Patients presented partially with hypothyroid (intellectual disability, motor developmental delay, brain atrophy, and constipation) and partially with hyperthyroid symptoms (tachycardia and behavioral abnormalities) to varying degrees. Functional characterization of THRA1p.(E173G) by reporter gene assays revealed increased transcriptional activity in contrast to THRA1(WT), unexpectedly revealing the first gain-of-function mutation found in THRA1. The THRA2 isoform does not bind T3 and antagonizes THRA1 action. Introduction of p.(E173G) into THRA2 increased its inhibitory effect on THRA1, which helps to explain the hypothyroid symptoms seen in our patients. We used protein structure models to investigate possible underlying pathomechanisms of this variant with a gain-of-antagonistic function and suggest that the p.(E173G) variant may have an influence on the dimerization domain of the nuclear receptor.
Keywords: gain-of-antagonistic function; gain-of-function; resistance to thyroid hormones; thyroid hormone receptor alpha.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Bichromatic Splicing Detector Allows Quantification of THRA1 and THRA2 Splicing Isoforms in Single Cells by Fluorescent Live-Cell Imaging.Int J Mol Sci. 2024 Dec 17;25(24):13512. doi: 10.3390/ijms252413512. Int J Mol Sci. 2024. PMID: 39769274 Free PMC article.
-
A Direct Comparison of Thyroid Hormone Receptor Protein Levels in Mice Provides Unexpected Insights into Thyroid Hormone Action.Thyroid. 2020 Aug;30(8):1193-1204. doi: 10.1089/thy.2019.0763. Epub 2020 Apr 6. Thyroid. 2020. PMID: 32122258 Free PMC article.
-
Thyroid hormone receptor mutations and disease: beyond thyroid hormone resistance.Trends Endocrinol Metab. 2005 May-Jun;16(4):176-82. doi: 10.1016/j.tem.2005.03.008. Trends Endocrinol Metab. 2005. PMID: 15860414 Review.
-
Thyroid hormone receptors and resistance to thyroid hormone disorders.Nat Rev Endocrinol. 2014 Oct;10(10):582-91. doi: 10.1038/nrendo.2014.143. Epub 2014 Aug 19. Nat Rev Endocrinol. 2014. PMID: 25135573 Free PMC article. Review.
-
Thyroid hormone receptor Thra and Thrb knockout differentially affects osteoblast biology and thyroid hormone responsiveness in vitro.J Cell Biochem. 2023 Dec;124(12):1948-1960. doi: 10.1002/jcb.30500. Epub 2023 Nov 22. J Cell Biochem. 2023. PMID: 37992217
Cited by
-
Dual Diagnosis of Nongoitrous Congenital Hypothyroidism-6 and Snijders Blok-Campeau Syndrome.Mol Syndromol. 2024 Jun;15(3):240-246. doi: 10.1159/000535682. Epub 2024 Jan 24. Mol Syndromol. 2024. PMID: 38841327 Free PMC article.
-
Clinical and Biochemical Characteristics of Untreated Adult Patients With Resistance to Thyroid Hormone Alpha.J Endocr Soc. 2023 Jul 4;7(8):bvad089. doi: 10.1210/jendso/bvad089. eCollection 2023 Jul 3. J Endocr Soc. 2023. PMID: 37469961 Free PMC article.
-
Mediator subunit MED1 differentially modulates mutant thyroid hormone receptor intracellular dynamics in Resistance to Thyroid Hormone syndrome.Mol Cell Endocrinol. 2023 Jan 1;559:111781. doi: 10.1016/j.mce.2022.111781. Epub 2022 Oct 1. Mol Cell Endocrinol. 2023. PMID: 36191835 Free PMC article.
-
Clinical and functional heterogeneity associated with the disruption of retinoic acid receptor beta.Genet Med. 2023 Aug;25(8):100856. doi: 10.1016/j.gim.2023.100856. Epub 2023 Apr 20. Genet Med. 2023. PMID: 37092537 Free PMC article.
-
Bichromatic Splicing Detector Allows Quantification of THRA1 and THRA2 Splicing Isoforms in Single Cells by Fluorescent Live-Cell Imaging.Int J Mol Sci. 2024 Dec 17;25(24):13512. doi: 10.3390/ijms252413512. Int J Mol Sci. 2024. PMID: 39769274 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
