

Modulation of the Gut Microbiota Structure with Probiotics and Isoflavone Alleviates Metabolic Disorder in Ovariectomized Mice
- PMID: 34070274
- PMCID: PMC8225012
- DOI: 10.3390/nu13061793
Modulation of the Gut Microbiota Structure with Probiotics and Isoflavone Alleviates Metabolic Disorder in Ovariectomized Mice
Abstract
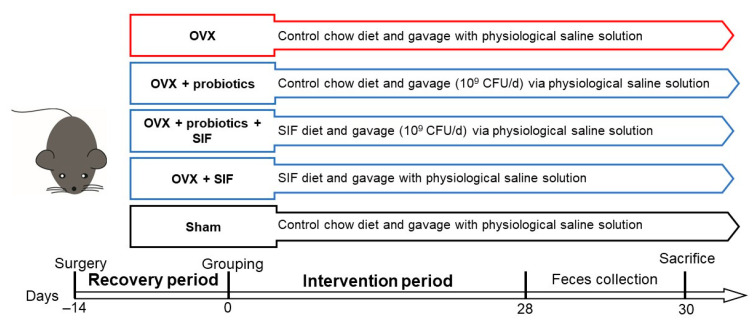
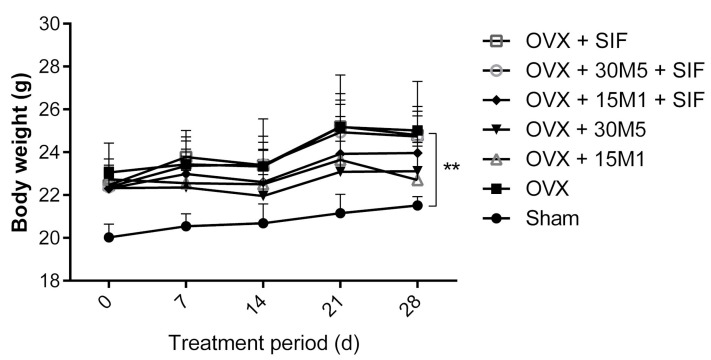
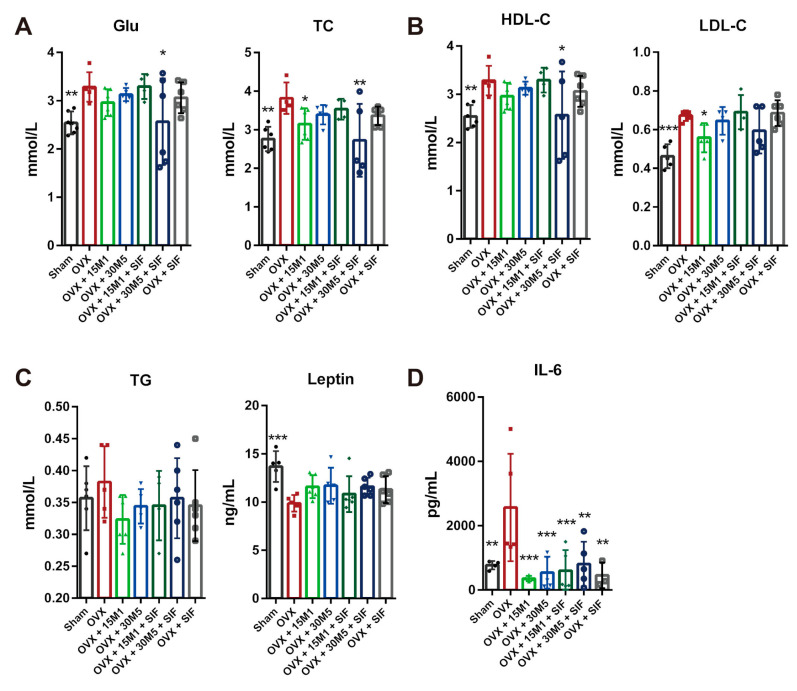
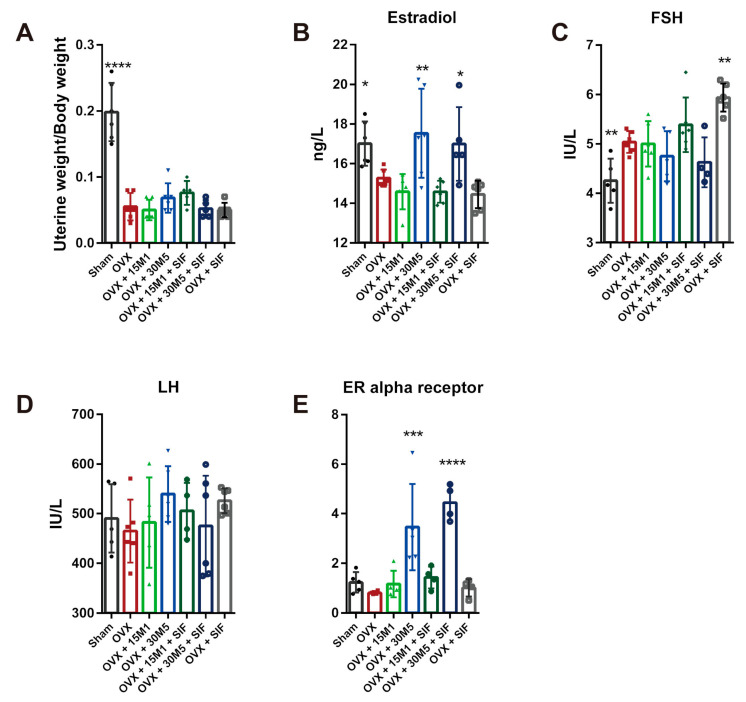
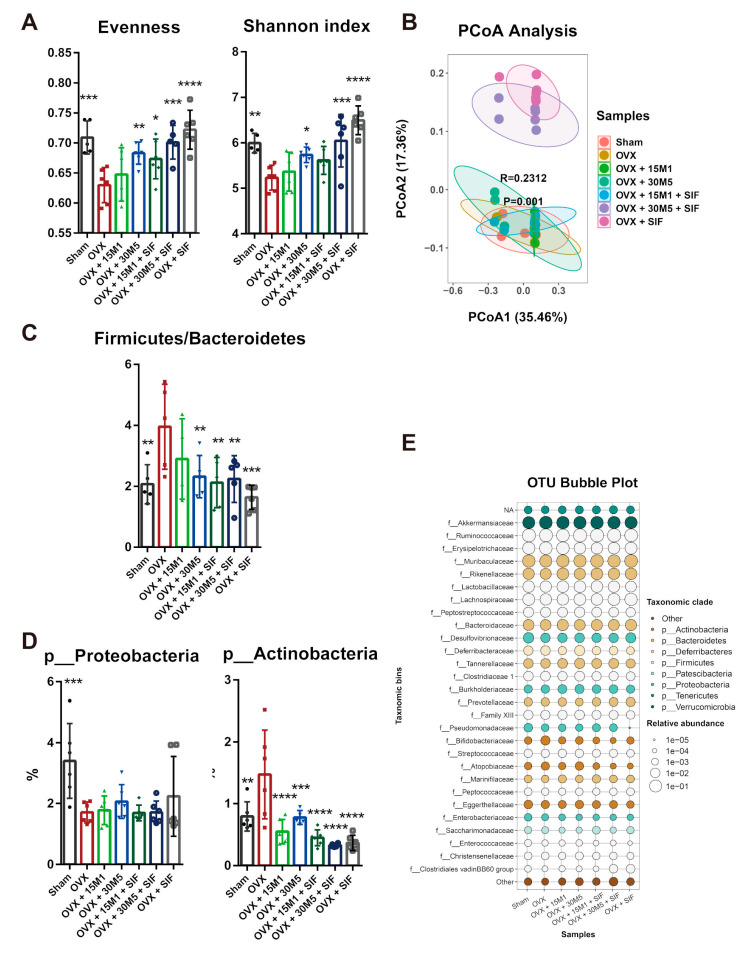
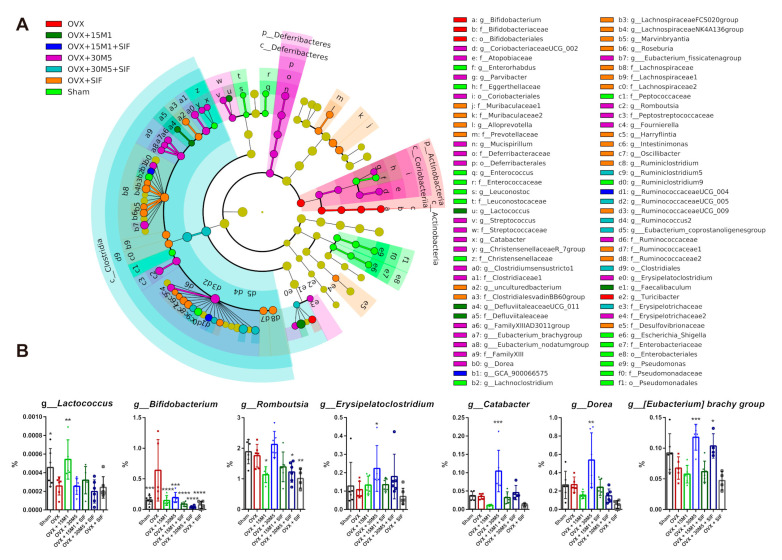
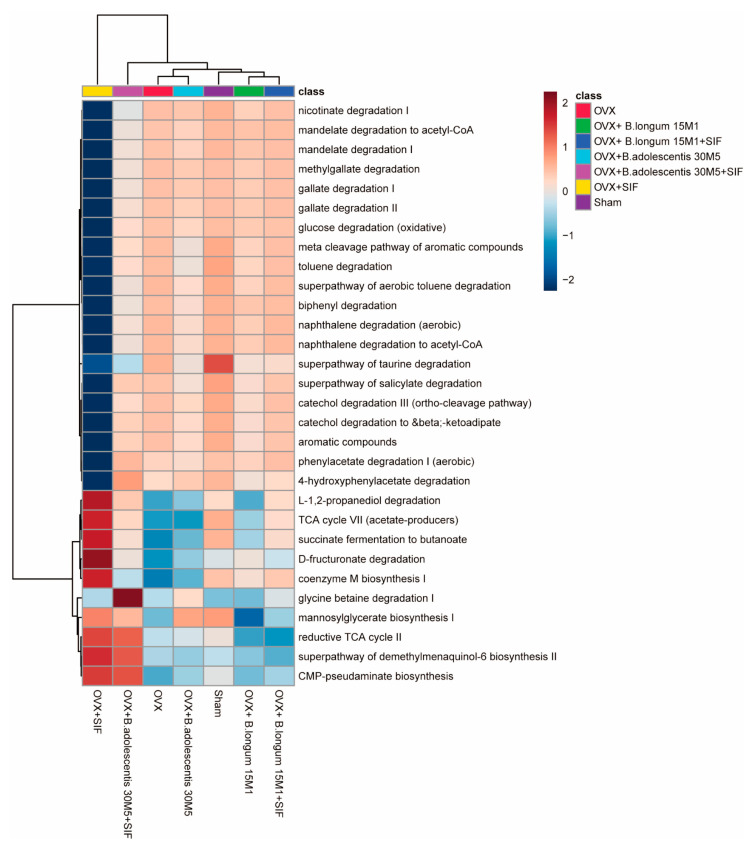
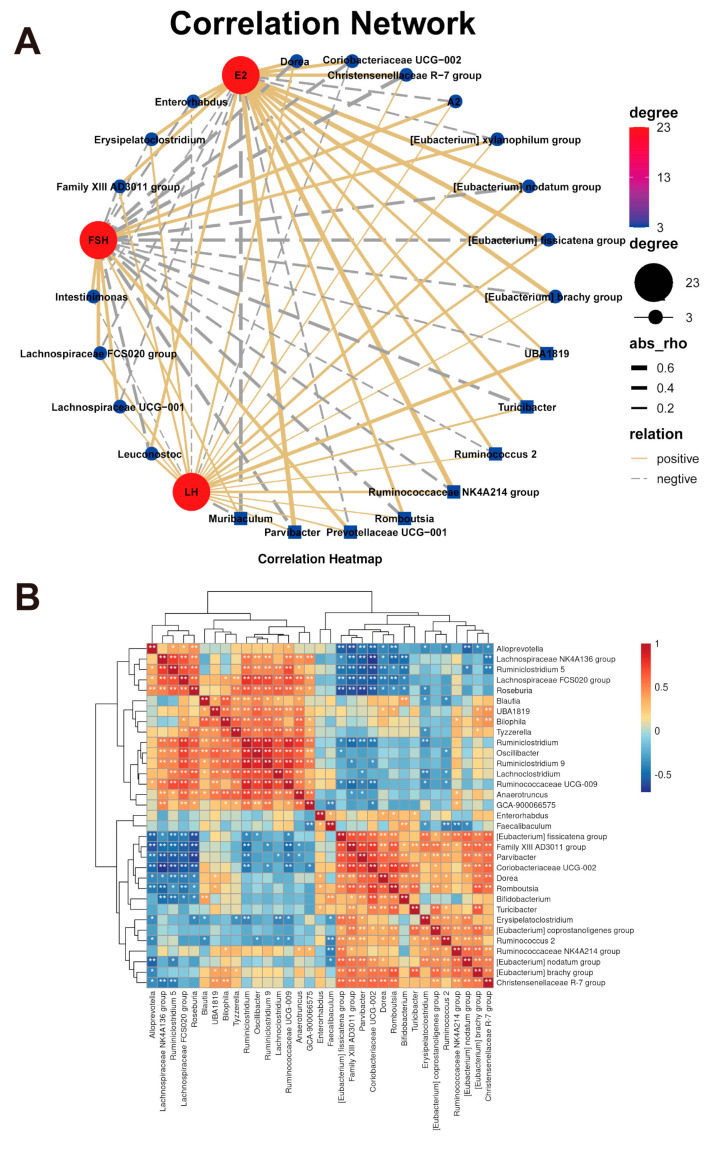
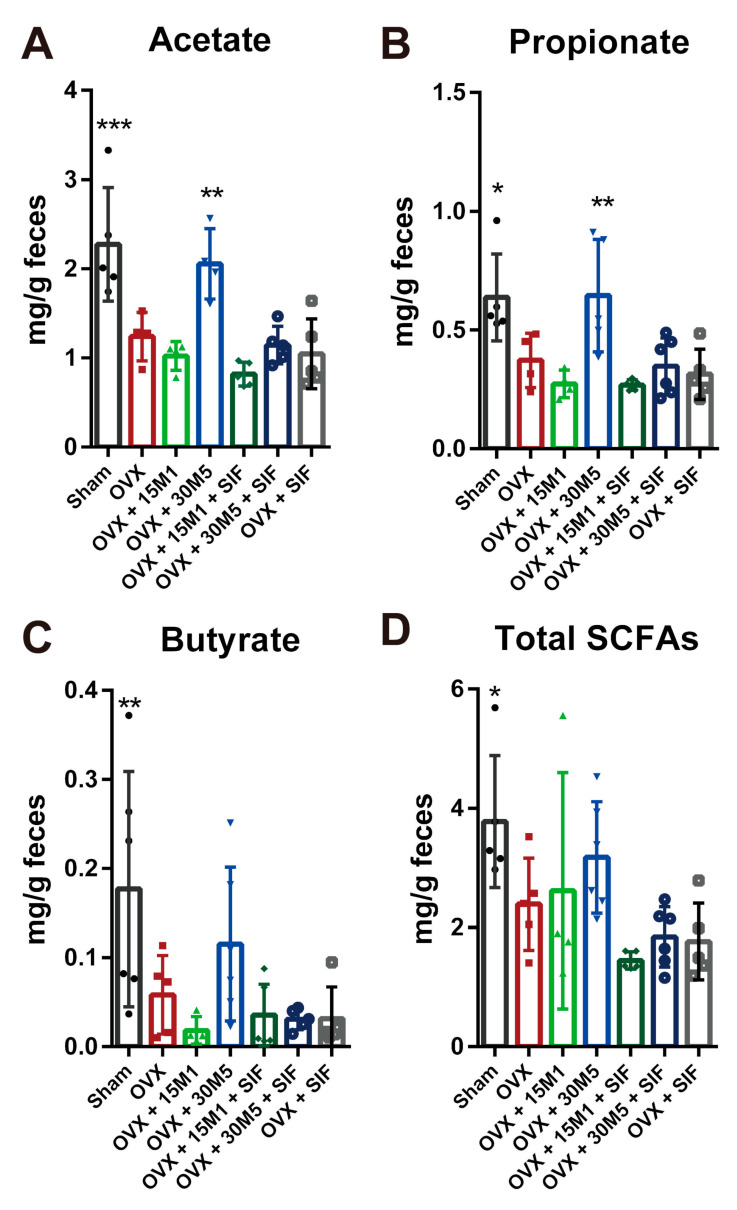
The decrease in ovarian hormone secretion that occurs during menopause results in an increase in body weight and adipose tissue mass. Probiotics and soy isoflavones (SIFs) could affect the gut microbiota and exert anti-obesity effects. The objective of this study was to investigate the effects of probiotics and a diet containing SIF (SIF diet) on ovariectomized mice with menopausal obesity, including the gut microbiome. The results demonstrate that Bifidobacterium longum 15M1 can reverse menopausal obesity, whilst the combination of Lactobacillus plantarum 30M5 and a SIF diet was more effective in alleviating menopausal lipid metabolism disorder than either components alone. Probiotics and SIFs play different anti-obesity roles in menopausal mice. Furthermore, 30M5 alters the metabolites of the gut microbiota that increase the circulating estrogen level, upregulates the expression of estrogen receptor α in abdominal adipose tissue and improves the production of short-chain fatty acids (SCFAs). A SIF diet can significantly alter the structure of the fecal bacterial community and enrich the pathways related to SCFAs production. Moreover, 30M5 and a SIF diet acted synergistically to effectively resolve abnormal serum lipid levels in ovariectomized mice, and these effects appear to be associated with regulation of the diversity and structure of the intestinal microbiota to enhance SCFAs production and promote estrogen circulation.
Keywords: estrogen; isoflavones; menopause; microbiota; obesity; probiotics; short-chain fatty acids.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Lactobacillus strains derived from human gut ameliorate metabolic disorders via modulation of gut microbiota composition and short-chain fatty acids metabolism.Benef Microbes. 2021 Jun 15;12(3):267-281. doi: 10.3920/BM2020.0148. Epub 2021 Jun 10. Benef Microbes. 2021. PMID: 34109894
-
Bifidobacterium adolescentis Isolated from Different Hosts Modifies the Intestinal Microbiota and Displays Differential Metabolic and Immunomodulatory Properties in Mice Fed a High-Fat Diet.Nutrients. 2021 Mar 21;13(3):1017. doi: 10.3390/nu13031017. Nutrients. 2021. PMID: 33801119 Free PMC article.
-
Anti-obesity effect of Lactobacillus rhamnosus LS-8 and Lactobacillus crustorum MN047 on high-fat and high-fructose diet mice base on inflammatory response alleviation and gut microbiota regulation.Eur J Nutr. 2020 Sep;59(6):2709-2728. doi: 10.1007/s00394-019-02117-y. Epub 2019 Oct 28. Eur J Nutr. 2020. PMID: 31659451
-
The Effect of Probiotics on the Production of Short-Chain Fatty Acids by Human Intestinal Microbiome.Nutrients. 2020 Apr 16;12(4):1107. doi: 10.3390/nu12041107. Nutrients. 2020. PMID: 32316181 Free PMC article. Review.
-
The Impact of Fermented Milk Products on Gut Microbiota-Derived Metabolites in Obesity: A Narrative Review.J Food Sci. 2025 Jun;90(6):e70301. doi: 10.1111/1750-3841.70301. J Food Sci. 2025. PMID: 40476741 Review.
Cited by
-
Microorganisms in the reproductive system and probiotic's regulatory effects on reproductive health.Comput Struct Biotechnol J. 2022 Mar 30;20:1541-1553. doi: 10.1016/j.csbj.2022.03.017. eCollection 2022. Comput Struct Biotechnol J. 2022. PMID: 35465162 Free PMC article. Review.
-
Impact of Probiotics and Prebiotics on Gut Microbiome and Hormonal Regulation.Gastrointest Disord (Basel). 2024 Dec;6(4):801-815. doi: 10.3390/gidisord6040056. Epub 2024 Sep 27. Gastrointest Disord (Basel). 2024. PMID: 39649015 Free PMC article.
-
The Anti-Inflammatory and Curative Exponent of Probiotics: A Comprehensive and Authentic Ingredient for the Sustained Functioning of Major Human Organs.Nutrients. 2024 Feb 16;16(4):546. doi: 10.3390/nu16040546. Nutrients. 2024. PMID: 38398870 Free PMC article. Review.
-
Heat-killed Limosilactobacillus reuteri ATCC PTA 6475 prevents bone loss in ovariectomized mice: A preliminary study.PLoS One. 2024 May 31;19(5):e0304358. doi: 10.1371/journal.pone.0304358. eCollection 2024. PLoS One. 2024. PMID: 38820403 Free PMC article.
-
Utilizing Lactic Acid Bacteria to Improve Hyperlipidemia: A Comprehensive Analysis from Gut Microbiota to Metabolic Pathways.Foods. 2024 Dec 16;13(24):4058. doi: 10.3390/foods13244058. Foods. 2024. PMID: 39767000 Free PMC article.
References
-
- Lee A., Ness R.B., Roman L.D., Terry K.L., Schildkraut J.M., Chang-Claude J., Doherty J.A., Menon U., Cramer D.W., Gayther S.A., et al. Association Between Menopausal Estrogen-Only Therapy and Ovarian Carcinoma Risk. Obstet. Gynecol. 2016;127:828–836. doi: 10.1097/AOG.0000000000001387. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
- No. 31972052, 32021005, 31820103010/the National Natural Science Foundation of China
- JUSRP22006, JUSRP51501/the Fundamental Research Funds for the Central Universities
- JUFSTR20180102/the national first-class discipline program of Food Science and Technology
- none/the Program of Collaborative Innovation Centre of Food Safety and Quality Control in Jiangsu Province
LinkOut - more resources
Full Text Sources
Medical
