

Novel S. cerevisiae Hybrid Synthetic Promoters Based on Foreign Core Promoter Sequences
- PMID: 34071849
- PMCID: PMC8198421
- DOI: 10.3390/ijms22115704
Novel S. cerevisiae Hybrid Synthetic Promoters Based on Foreign Core Promoter Sequences
Abstract
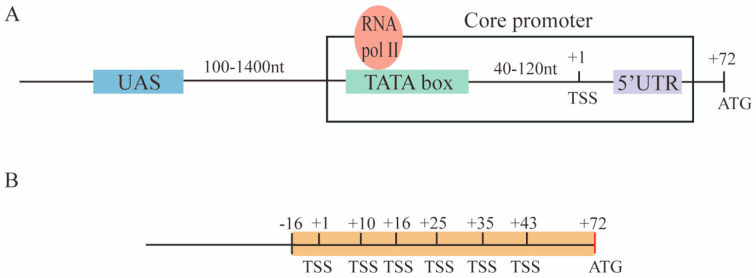
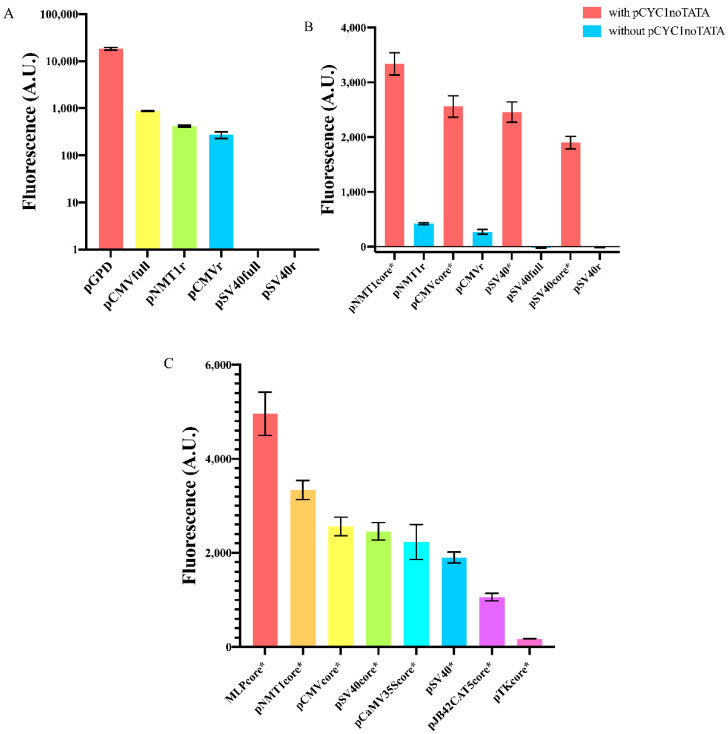
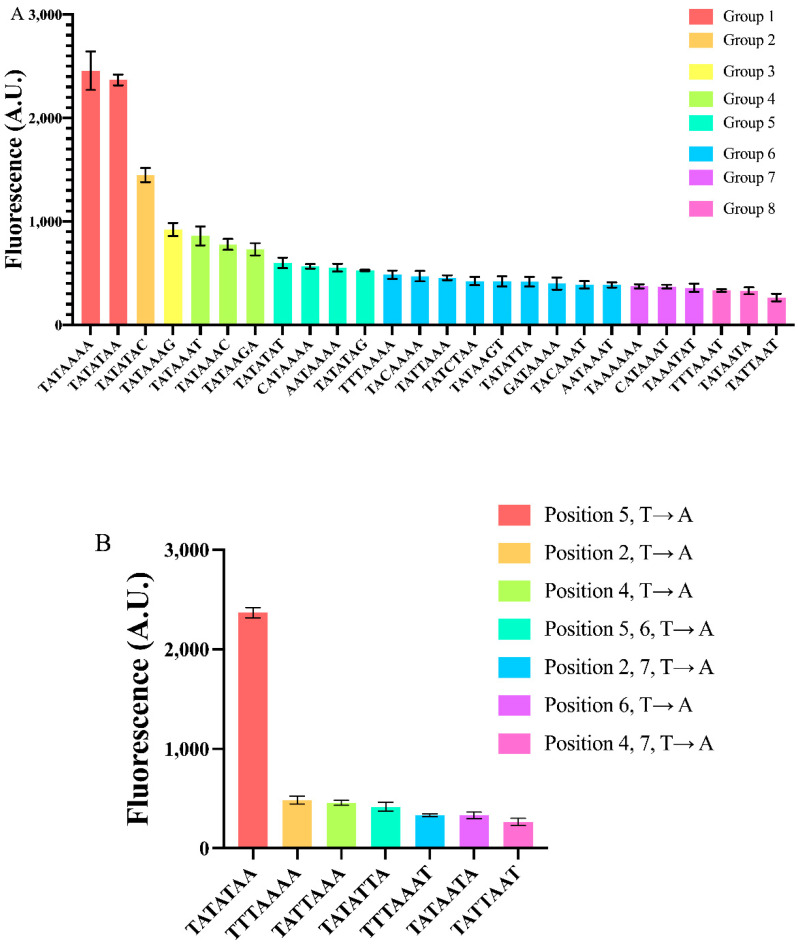
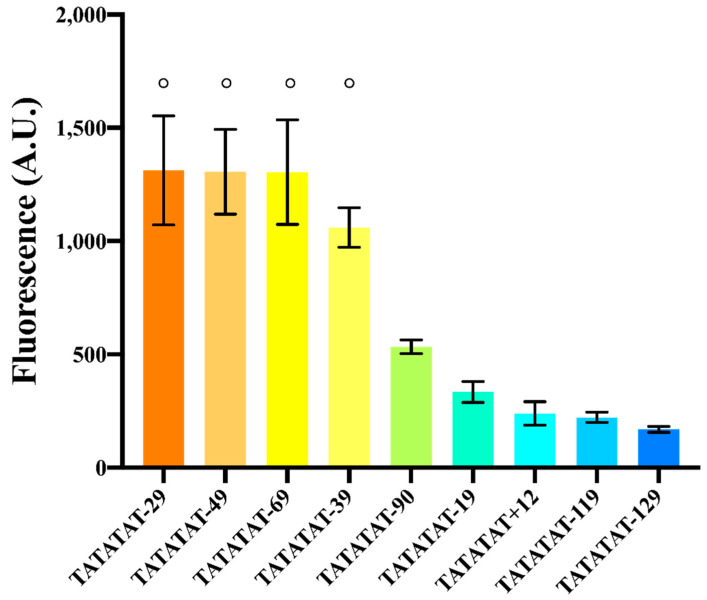
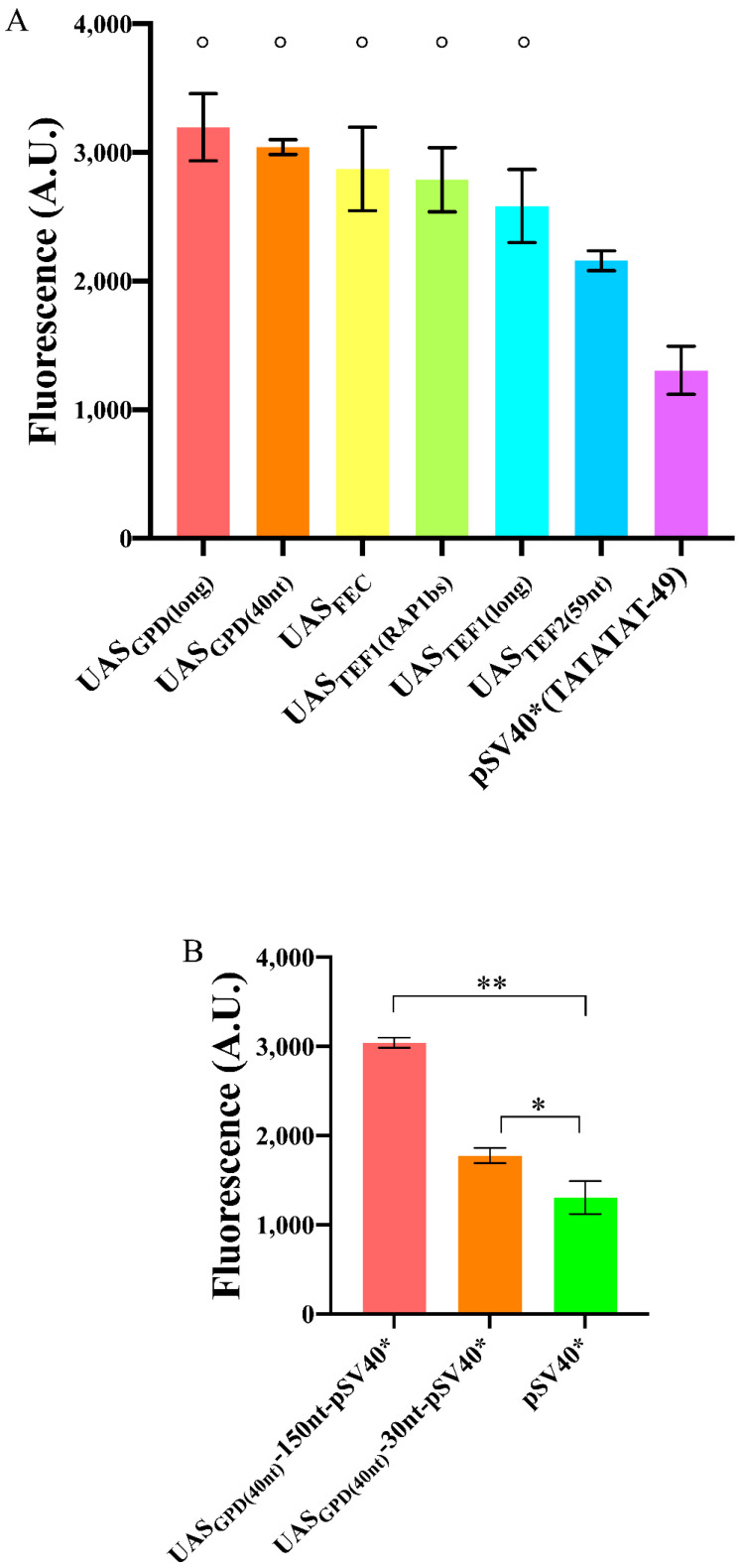
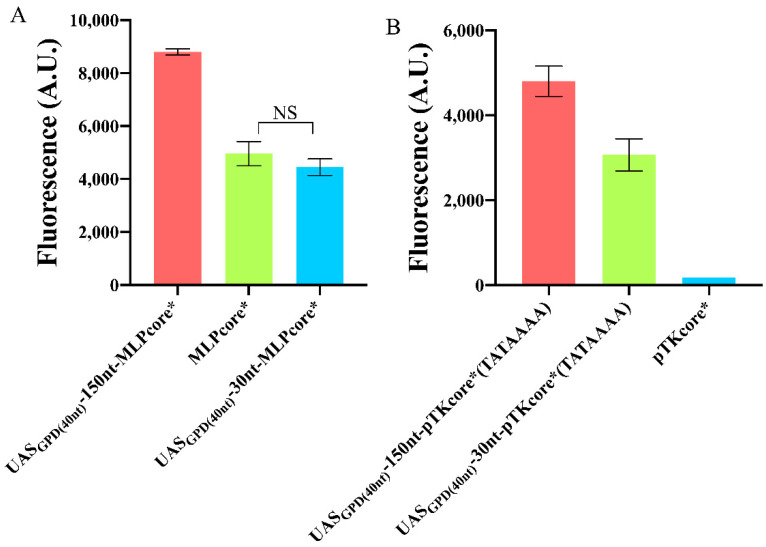
Promoters are fundamental components of synthetic gene circuits. They are DNA segments where transcription initiation takes place. New constitutive and regulated promoters are constantly engineered in order to meet the requirements for protein and RNA expression into different genetic networks. In this work, we constructed and optimized new synthetic constitutive promoters for the yeast Saccharomyces cerevisiae. We started from foreign (e.g., viral) core promoters as templates. They are, usually, unfunctional in yeast but can be activated by extending them with a short sequence, from the CYC1 promoter, containing various transcription start sites (TSSs). Transcription was modulated by mutating the TATA box composition and varying its distance from the TSS. We found that gene expression is maximized when the TATA box has the form TATAAAA or TATATAA and lies between 30 and 70 nucleotides upstream of the TSS. Core promoters were turned into stronger promoters via the addition of a short UAS. In particular, the 40 nt bipartite UAS from the GPD promoter can enhance protein synthesis considerably when placed 150 nt upstream of the TATA box. Overall, we extended the pool of S. cerevisiae promoters with 59 new samples, the strongest overcoming the native TEF2 promoter.
Keywords: Saccharomyces cerevisiae; TATA box; UAS; gene expression; promoters; synthetic biology.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
A Random Screen Using a Novel Reporter Assay System Reveals a Set of Sequences That Are Preferred as the TATA or TATA-Like Elements in the CYC1 Promoter of Saccharomyces cerevisiae.PLoS One. 2015 Jun 5;10(6):e0129357. doi: 10.1371/journal.pone.0129357. eCollection 2015. PLoS One. 2015. PMID: 26046838 Free PMC article.
-
SAGA mediates transcription from the TATA-like element independently of Taf1p/TFIID but dependent on core promoter structures in Saccharomyces cerevisiae.PLoS One. 2017 Nov 27;12(11):e0188435. doi: 10.1371/journal.pone.0188435. eCollection 2017. PLoS One. 2017. PMID: 29176831 Free PMC article.
-
A conserved GA element in TATA-less RNA polymerase II promoters.PLoS One. 2011;6(11):e27595. doi: 10.1371/journal.pone.0027595. Epub 2011 Nov 16. PLoS One. 2011. PMID: 22110682 Free PMC article.
-
The pattern and evolution of yeast promoter bendability.Trends Genet. 2007 Jul;23(7):318-21. doi: 10.1016/j.tig.2007.03.015. Epub 2007 Apr 6. Trends Genet. 2007. PMID: 17418911 Review.
-
Biological functions of the duplicated GGAA-motifs in various human promoter regions.Yakugaku Zasshi. 2011;131(12):1787-800. doi: 10.1248/yakushi.131.1787. Yakugaku Zasshi. 2011. PMID: 22129877 Review.
Cited by
-
Natural promoters and promoter engineering strategies for metabolic regulation in Saccharomyces cerevisiae.J Ind Microbiol Biotechnol. 2023 Feb 17;50(1):kuac029. doi: 10.1093/jimb/kuac029. J Ind Microbiol Biotechnol. 2023. PMID: 36633543 Free PMC article. Review.
-
Hybrid Boolean gates show that Cas12c controls transcription activation effectively in the yeast S. cerevisiae.Front Bioeng Biotechnol. 2023 Sep 12;11:1267174. doi: 10.3389/fbioe.2023.1267174. eCollection 2023. Front Bioeng Biotechnol. 2023. PMID: 37771576 Free PMC article.
-
Hybrid Synthetic Promoters in Saccharomyces cerevisiae Built on Foreign Promoter Sequences.Methods Mol Biol. 2024;2844:109-119. doi: 10.1007/978-1-0716-4063-0_7. Methods Mol Biol. 2024. PMID: 39068335
-
Synthetic polycistronic sequences in eukaryotes.Synth Syst Biotechnol. 2021 Sep 15;6(4):254-261. doi: 10.1016/j.synbio.2021.09.003. eCollection 2021 Dec. Synth Syst Biotechnol. 2021. PMID: 34584993 Free PMC article. Review.
-
Engineering of Bacillus Promoters Based on Interacting Motifs between UP Elements and RNA Polymerase (RNAP) α-Subunit.Int J Mol Sci. 2022 Nov 3;23(21):13480. doi: 10.3390/ijms232113480. Int J Mol Sci. 2022. PMID: 36362266 Free PMC article.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
