

LncRNA PART1 Promotes Proliferation and Migration, Is Associated with Cancer Stem Cells, and Alters the miRNA Landscape in Triple-Negative Breast Cancer
- PMID: 34072264
- PMCID: PMC8198907
- DOI: 10.3390/cancers13112644
LncRNA PART1 Promotes Proliferation and Migration, Is Associated with Cancer Stem Cells, and Alters the miRNA Landscape in Triple-Negative Breast Cancer
Abstract
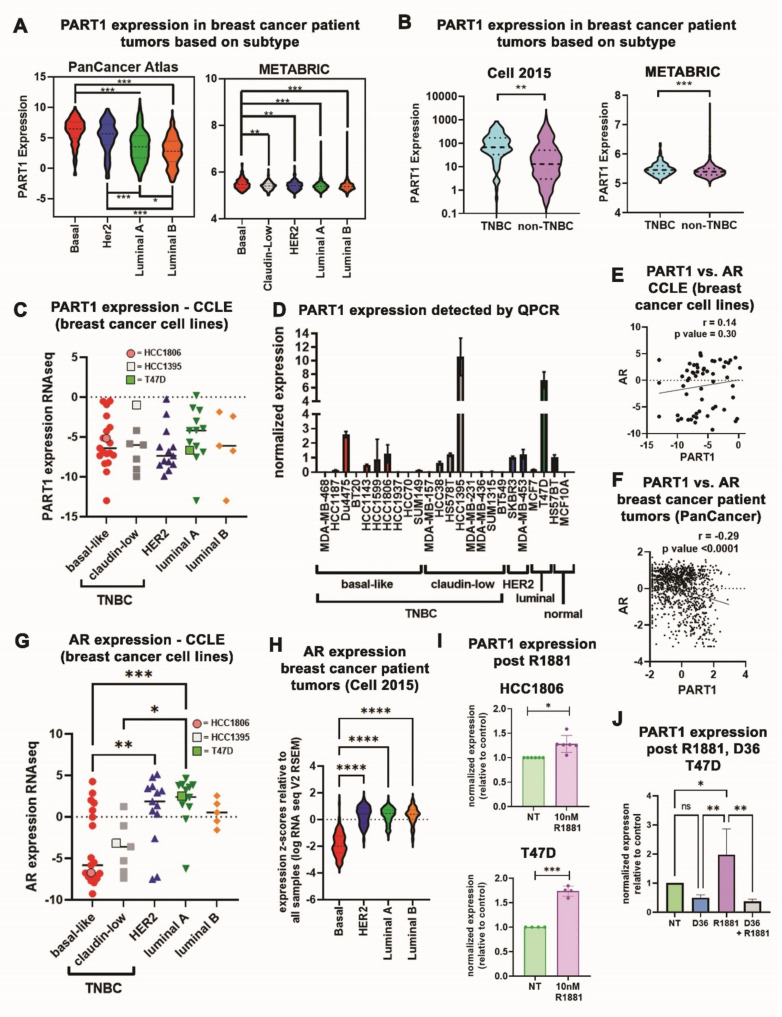
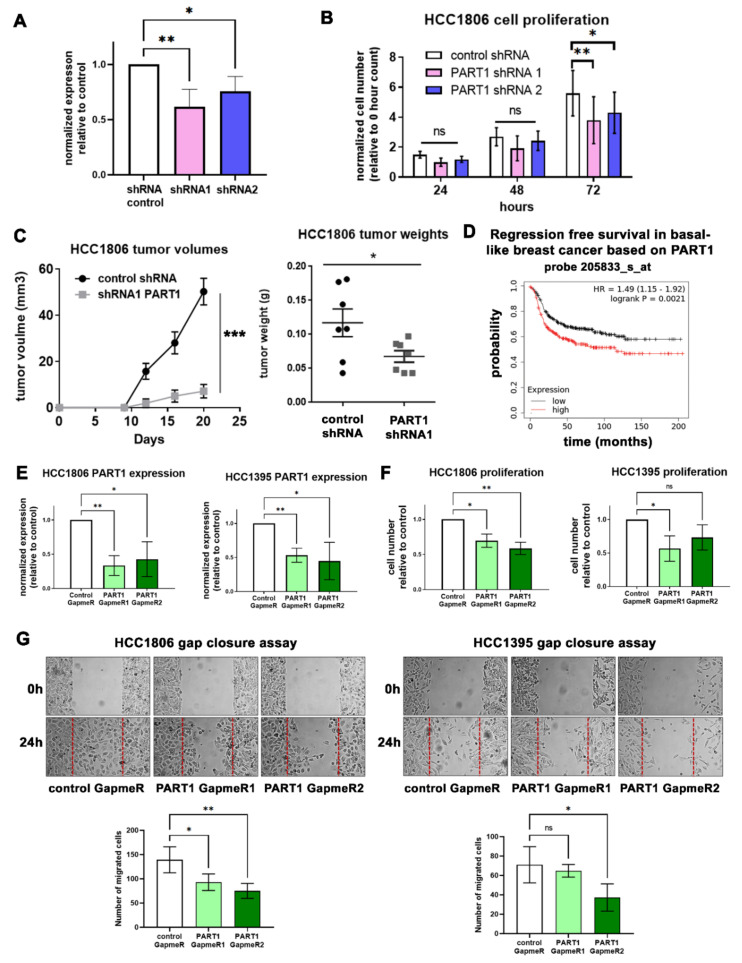
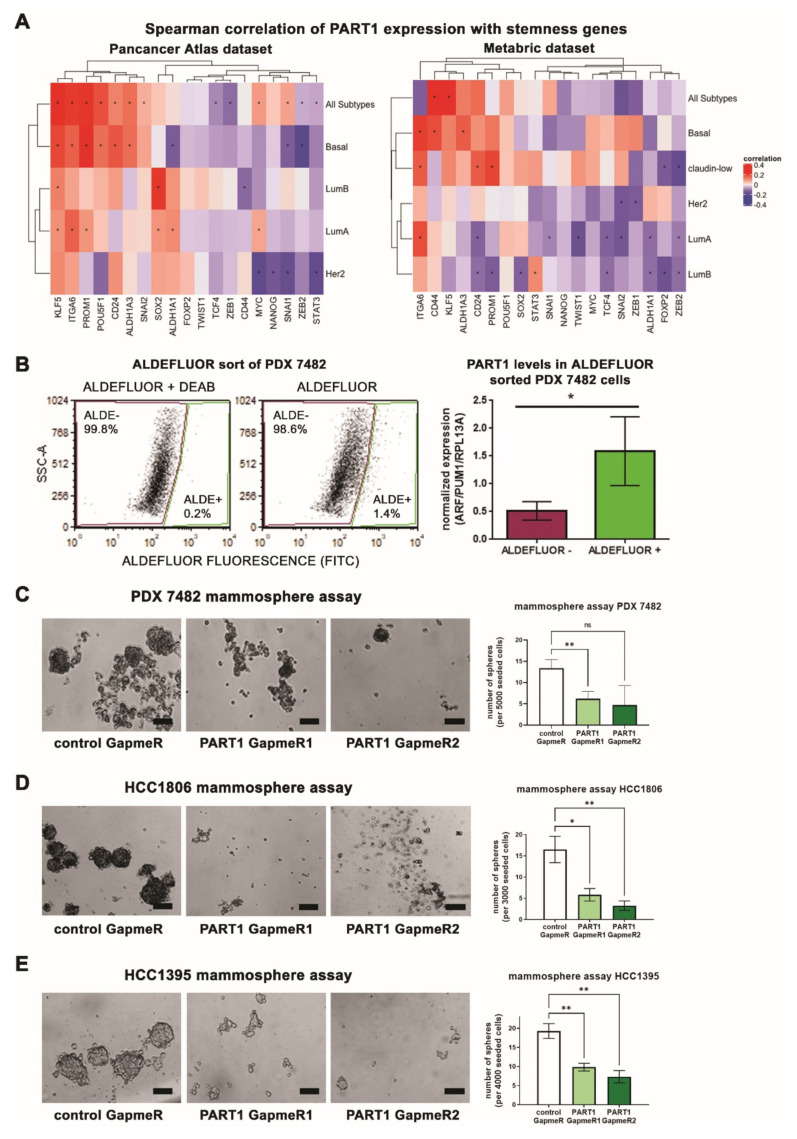
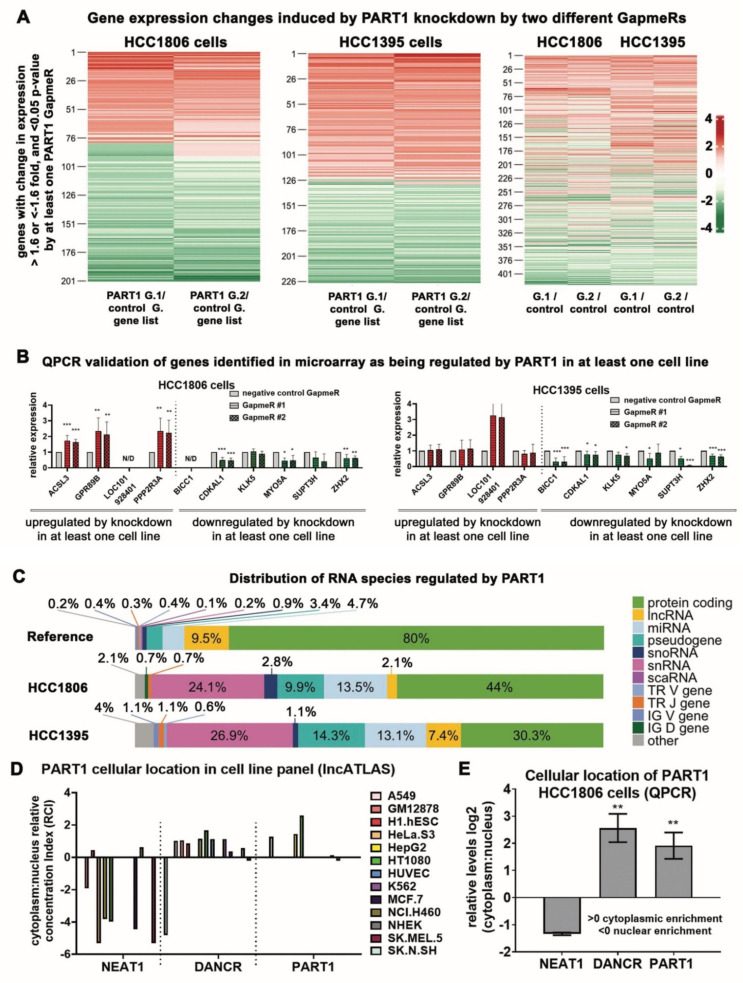
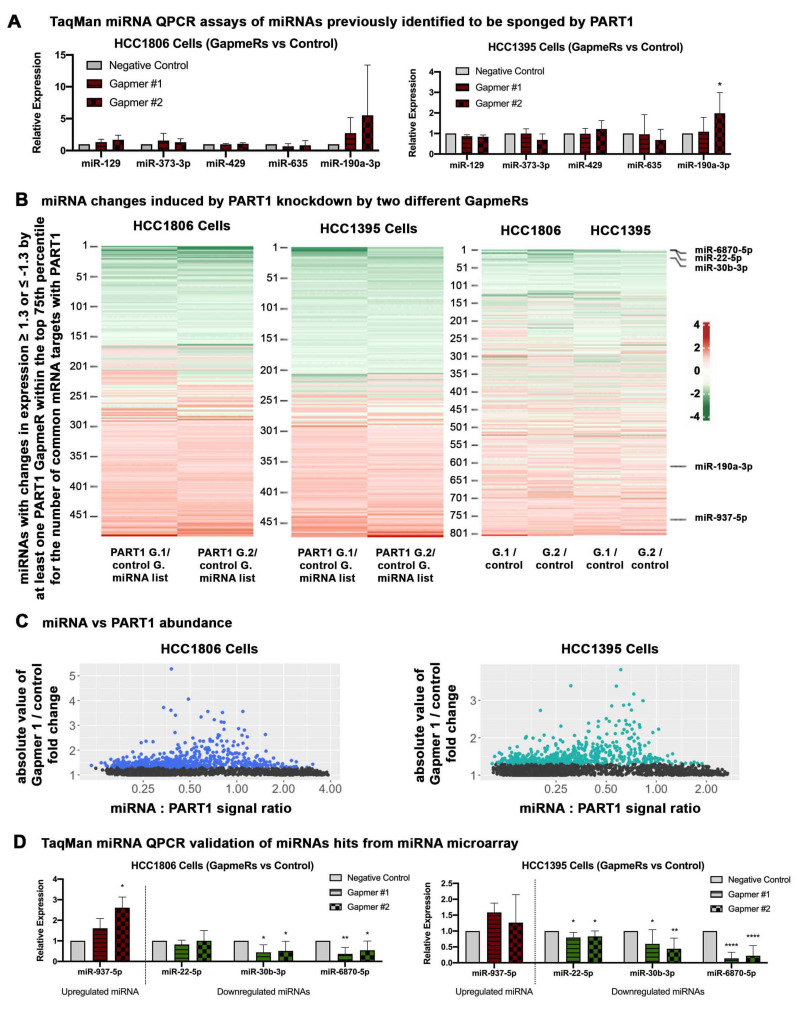
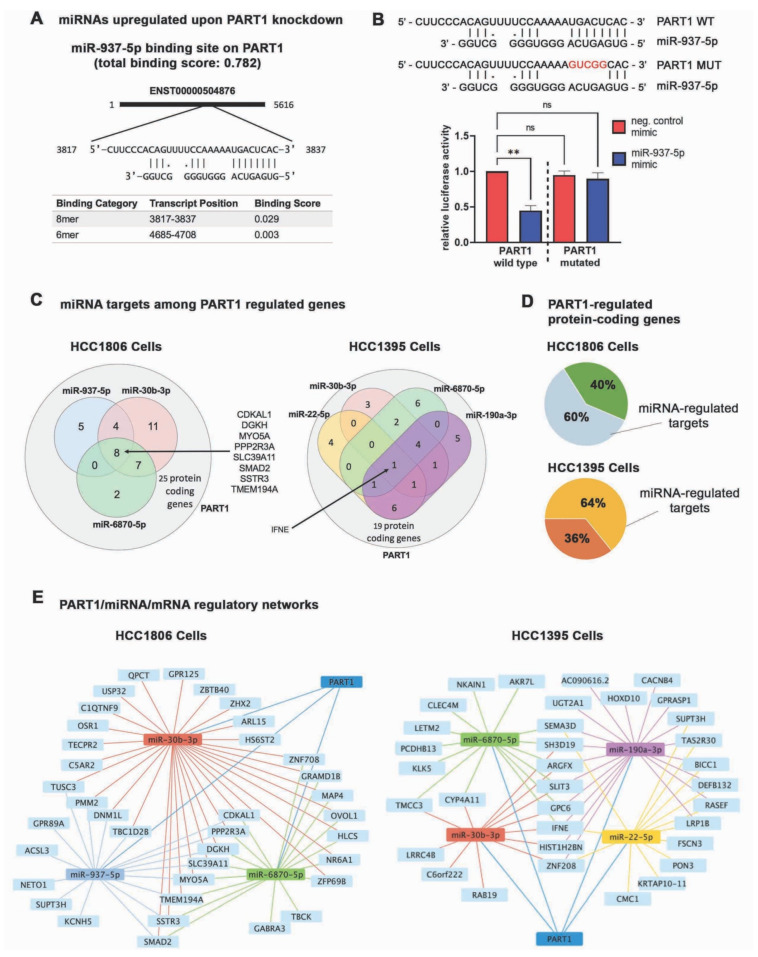
Triple-negative breast cancers (TNBCs) are aggressive, lack targeted therapies and are enriched in cancer stem cells (CSCs). Novel therapies which target CSCs within these tumors would likely lead to improved outcomes for TNBC patients. Long non-coding RNAs (lncRNAs) are potential therapeutic targets for TNBC and CSCs. We demonstrate that lncRNA prostate androgen regulated transcript 1 (PART1) is enriched in TNBCs and in Aldefluorhigh CSCs, and is associated with worse outcomes among basal-like breast cancer patients. Although PART1 is androgen inducible in breast cancer cells, analysis of patient tumors indicates its androgen regulation has minimal clinical impact. Knockdown of PART1 in TNBC cell lines and a patient-derived xenograft decreased cell proliferation, migration, tumor growth, and mammosphere formation potential. Transcriptome analyses revealed that the lncRNA affects expression of hundreds of genes (e.g., myosin-Va, MYO5A; zinc fingers and homeoboxes protein 2, ZHX2). MiRNA 4.0 GeneChip and TaqMan assays identified multiple miRNAs that are regulated by cytoplasmic PART1, including miR-190a-3p, miR-937-5p, miR-22-5p, miR-30b-3p, and miR-6870-5p. We confirmed the novel interaction between PART1 and miR-937-5p. In general, miRNAs altered by PART1 were less abundant than PART1, potentially leading to cell line-specific effects in terms miRNA-PART1 interactions and gene regulation. Together, the altered miRNA landscape induced by PART1 explains most of the protein-coding gene regulation changes (e.g., MYO5A) induced by PART1 in TNBC.
Keywords: CSCs; PART1; TNBC; lncRNA; miRNAs.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Pandya G., Kirtonia A., Sethi G., Pandey A.K., Garg M. The implication of long non-coding RNAs in the diagnosis, pathogenesis and drug resistance of pancreatic ductal adenocarcinoma and their possible therapeutic potential. Biochim. Biophys. Acta Rev. Cancer. 2020;1874:188423. doi: 10.1016/j.bbcan.2020.188423. - DOI - PubMed
-
- Prensner J.R., Chinnaiyan A.M. The Emergence of lncRNAs in Cancer Biology. Cancer Discov. 2011;1:391–407. doi: 10.1158/2159-8290.CD-11-0209. - DOI - PMC - PubMed
-
- Vidovic D., Huynh T.T., Konda P., Dean C., Cruickshank B.M., Sultan M., Coyle K.M., Gujar S., Marcato P. ALDH1A3-regulated long non-coding RNA NRAD1 is a potential novel target for triple-negative breast tumors and cancer stem cells. Cell Death Differ. 2019 doi: 10.1038/s41418-019-0362-1. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
