

Optimization of Culture Conditions for Oxygen-Tolerant Regulatory [NiFe]-Hydrogenase Production from Ralstonia eutropha H16 in Escherichia coli
- PMID: 34073092
- PMCID: PMC8229454
- DOI: 10.3390/microorganisms9061195
Optimization of Culture Conditions for Oxygen-Tolerant Regulatory [NiFe]-Hydrogenase Production from Ralstonia eutropha H16 in Escherichia coli
Abstract
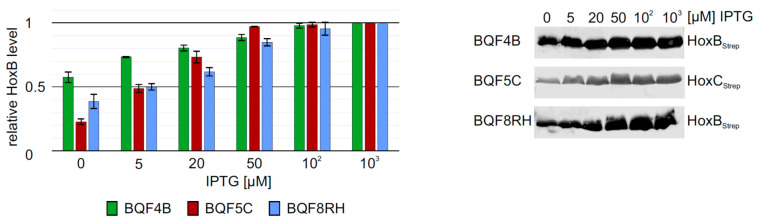
Hydrogenases are abundant metalloenzymes that catalyze the reversible conversion of molecular H2 into protons and electrons. Important achievements have been made over the past two decades in the understanding of these highly complex enzymes. However, most hydrogenases have low production yields requiring many efforts and high costs for cultivation limiting their investigation. Heterologous production of these hydrogenases in a robust and genetically tractable expression host is an attractive strategy to make these enzymes more accessible. In the present study, we chose the oxygen-tolerant H2-sensing regulatory [NiFe]-hydrogenase (RH) from Ralstonia eutropha H16 owing to its relatively simple architecture compared to other [NiFe]-hydrogenases as a model to develop a heterologous hydrogenase production system in Escherichia coli. Using screening experiments in 24 deep-well plates with 3 mL working volume, we investigated relevant cultivation parameters, including inducer concentration, expression temperature, and expression time. The RH yield could be increased from 14 mg/L up to >250 mg/L by switching from a batch to an EnPresso B-based fed-batch like cultivation in shake flasks. This yield exceeds the amount of RH purified from the homologous host R. eutropha by several 100-fold. Additionally, we report the successful overproduction of the RH single subunits HoxB and HoxC, suitable for biochemical and spectroscopic investigations. Even though both RH and HoxC proteins were isolated in an inactive, cofactor free apo-form, the proposed strategy may powerfully accelerate bioprocess development and structural studies for both basic research and applied studies. These results are discussed in the context of the regulation mechanisms governing the assembly of large and small hydrogenase subunits.
Keywords: Escherichia coli; Ralstonia eutropha; [NiFe]-hydrogenase; cofactor assembly; difficult-to-express protein; heterologous protein production.
Conflict of interest statement
The authors declare no conflict of interest.
Figures






References
-
- Greening C., Biswas A., Carere C.R., Jackson C.J., Taylor M.C., Stott M.B., Cook G.M., Morales S.E. Genomic and metagenomic surveys of hydrogenase distribution indicate H2 is a widely utilised energy source for microbial growth and survival. ISME J. 2016;10:761–777. doi: 10.1038/ismej.2015.153. - DOI - PMC - PubMed
-
- Preissler J., Reeve H.A., Zhu T., Nicholson J., Urata K., Lauterbach L., Wong L.L., Vincent K.A., Lenz O. Dihydrogen-Driven NADPH Recycling in Imine Reduction and P450-Catalyzed Oxidations Mediated by an Engineered O2-Tolerant Hydrogenase. ChemCatChem. 2020;12:4853–4861. doi: 10.1002/cctc.202000763. - DOI
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
