

Osmolyte Signatures for the Protection of Aspergillus sydowii Cells under Halophilic Conditions and Osmotic Shock
- PMID: 34073303
- PMCID: PMC8228332
- DOI: 10.3390/jof7060414
Osmolyte Signatures for the Protection of Aspergillus sydowii Cells under Halophilic Conditions and Osmotic Shock
Abstract
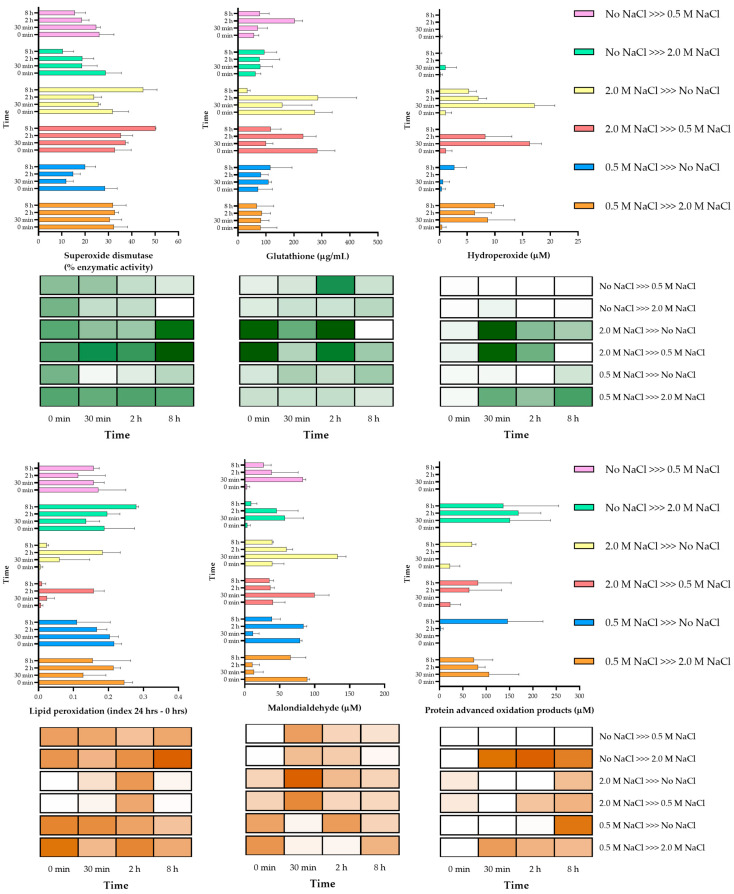
Aspergillus sydowii is a moderate halophile fungus extensively studied for its biotechnological potential and halophile responses, which has also been reported as a coral reef pathogen. In a recent publication, the transcriptomic analysis of this fungus, when growing on wheat straw, showed that genes related to cell wall modification and cation transporters were upregulated under hypersaline conditions but not under 0.5 M NaCl, the optimal salinity for growth in this strain. This led us to study osmolyte accumulation as a mechanism to withstand moderate salinity. In this work, we show that A. sydowii accumulates trehalose, arabitol, mannitol, and glycerol with different temporal dynamics, which depend on whether the fungus is exposed to hypo- or hyperosmotic stress. The transcripts coding for enzymes responsible for polyalcohol synthesis were regulated in a stress-dependent manner. Interestingly, A. sydowii contains three homologs (Hog1, Hog2 and MpkC) of the Hog1 MAPK, the master regulator of hyperosmotic stress response in S. cerevisiae and other fungi. We show a differential regulation of these MAPKs under different salinity conditions, including sustained basal Hog1/Hog2 phosphorylation levels in the absence of NaCl or in the presence of 2.0 M NaCl, in contrast to what is observed in S. cerevisiae. These findings indicate that halophilic fungi such as A. sydowii utilize different osmoadaptation mechanisms to hypersaline conditions.
Keywords: Aspergillus; HOG; extremophile; halophile; osmolyte; osmotic shock.
Conflict of interest statement
The authors declare no conflict of interest. Also, the funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures








References
-
- Brown A.D., Mackenzie K.F., Singh K.K. Selected aspects of microbial osmoregulation. FEMS Microbiol. Lett. 1986;39:31–36. doi: 10.1111/j.1574-6968.1986.tb01839.x. - DOI
Grants and funding
LinkOut - more resources
Full Text Sources
