

Review
doi: 10.1101/cshperspect.a040550.
NADPH-The Forgotten Reducing Equivalent
Affiliations
- PMID: 34074675
- PMCID: PMC8168431
- DOI: 10.1101/cshperspect.a040550
Item in Clipboard
Review
NADPH-The Forgotten Reducing Equivalent
Cold Spring Harb Perspect Biol.
.
No abstract available
Figures
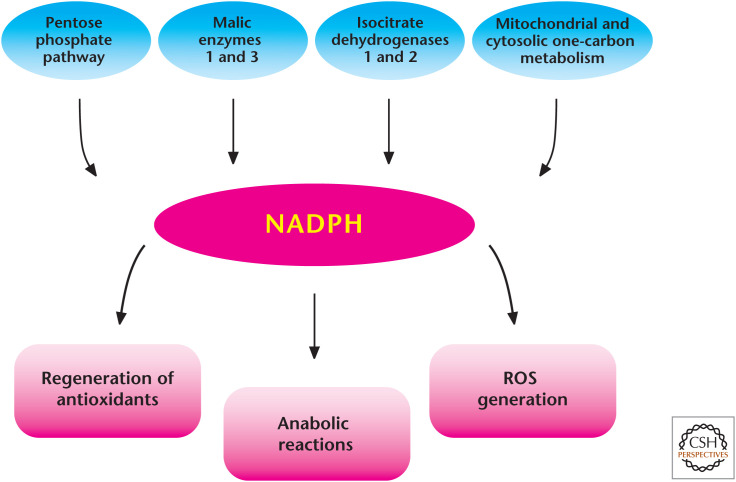
Diverse roles of NADPH in metabolism. There are multiple sources that generate NADPH in the mitochondria and cytosol. NADPH is critical for many anabolic reactions and is essential to maintain antioxidant capacity in cells. NADPH can also be used to generate ROS through NADPH oxidases.
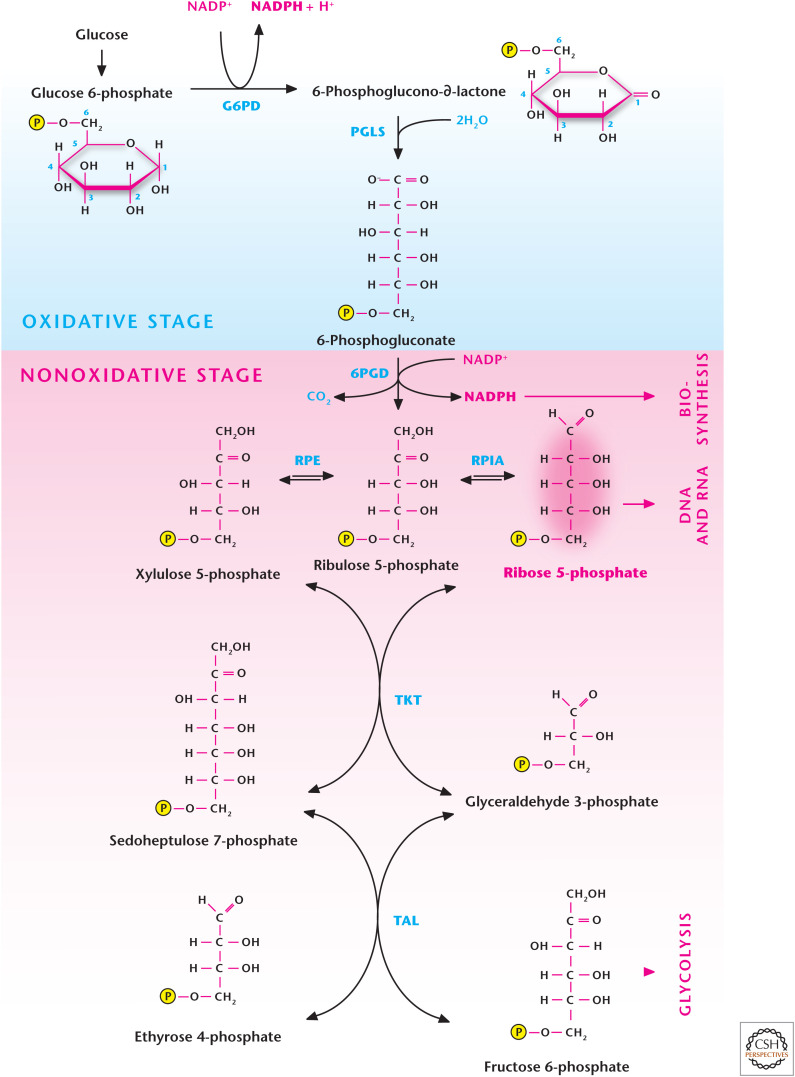
The pentose phosphate pathway (PPP). Glucose conversion to glucose 6-phosphate by G6PD in step 1 of glycolysis allows for the funneling of the latter metabolite down the glycolytic pathway or into the PPP. The oxidative stage of the PPP generates NADPH, whereas the nonoxidative stage generates ribose 5-phosphate, which can be used for DNA and RNA synthesis. G6PD, glucose 6-phosphate hydrogenase; PGLS, 6-phosphogluconolactonase; 6PGD, 6-phosphogluconate dehydrogenase; RPE, ribulose 5-phosphate-3-epimerase; RPIA, ribose 5-phosphate isomerase A; TKT, transketolase; TAL, transaldolase.
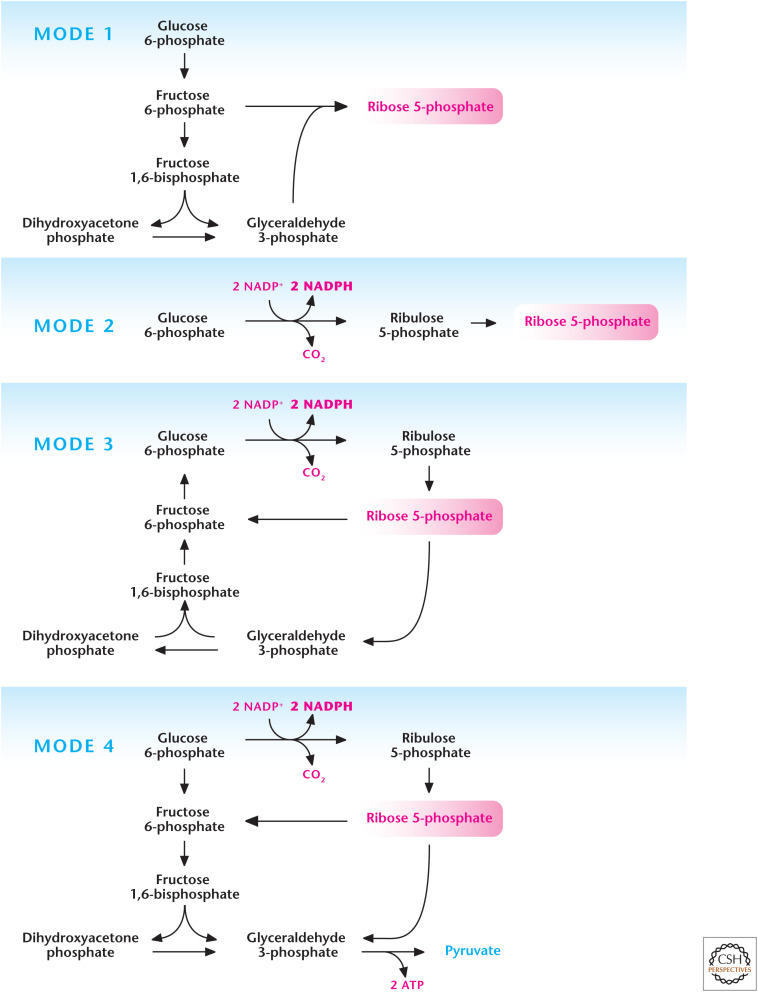
Different modes of the pentose phosphate pathway (PPP). The PPP can operate in different modes depending on cellular requirements of NADPH and ribose 5-phosphate. Mode 1 primarily generates ribose 5-phosphate by funneling glycolytic intermediates into the nonoxidative stage of PPP. Mode 2 produces balance of both NADPH and ribose 5-phosphate. Mode 3 produces primarily NADPH as the ribose 5-phosphate is funneled into glycolysis to go back into the oxidative stage of PPP for further generation of NADPH. Mode 4 generates NADPH and the ribose 5-phosphate is funneled into glycolysis to generate ATP. (Adapted, with permission, from Berg et al. 2012, © W.H. Freeman and Company.)
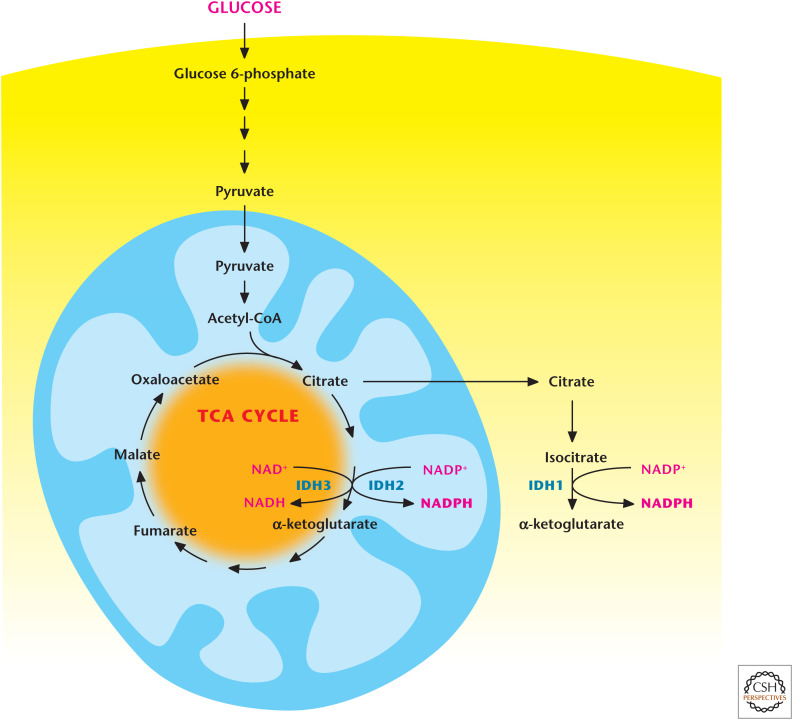
Isocitrate dehydrogenases 1 and 2 produce NAPDH. Cytosolic isocitrate dehydrogenase 1 (IDH1) and mitochondrial isocitrate dehydrogenase 2 (IDH2) produce NADPH through conversion of isocitrate into α-ketoglutarate. Note that mitochondrial isocitrate dehydrogenase 3 (IDH3) generates NADH by converting isocitrate into α-ketoglutarate.
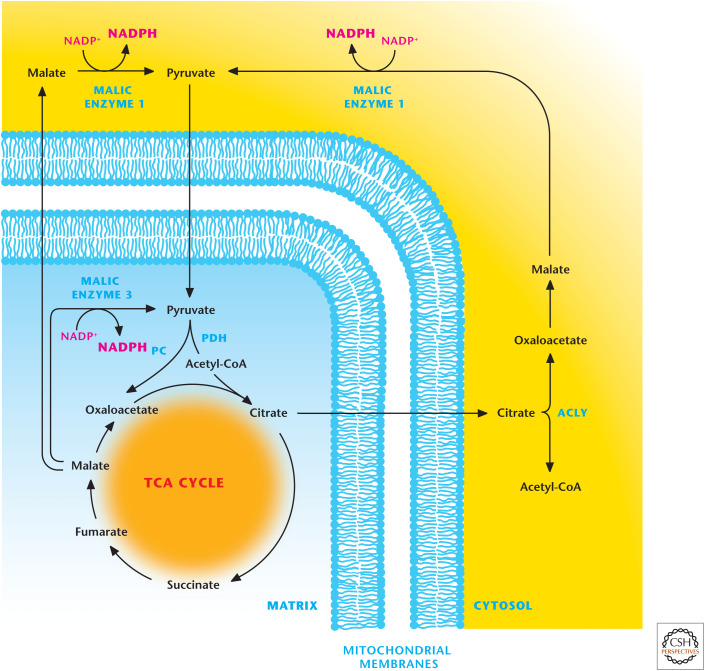
Malic enzymes 1 and 3 produce NADPH. Mitochondrial malic enzyme 3 can generate NADPH by converting malate into pyruvate. Malate can also exit from the TCA cycle where it is converted into pyruvate by malic enzyme 1, resulting in the production of NADPH. The TCA-cycle intermediate citrate can be transported into the cytosol where it is converted into acetyl-CoA and oxaloacetate by ATP-citrate lyase (ACLY). The latter is converted into malate to produce NADPH by malic enzyme 1. PC, pyruvate carboxylase; PDH, pyruvate dehydrogenase.
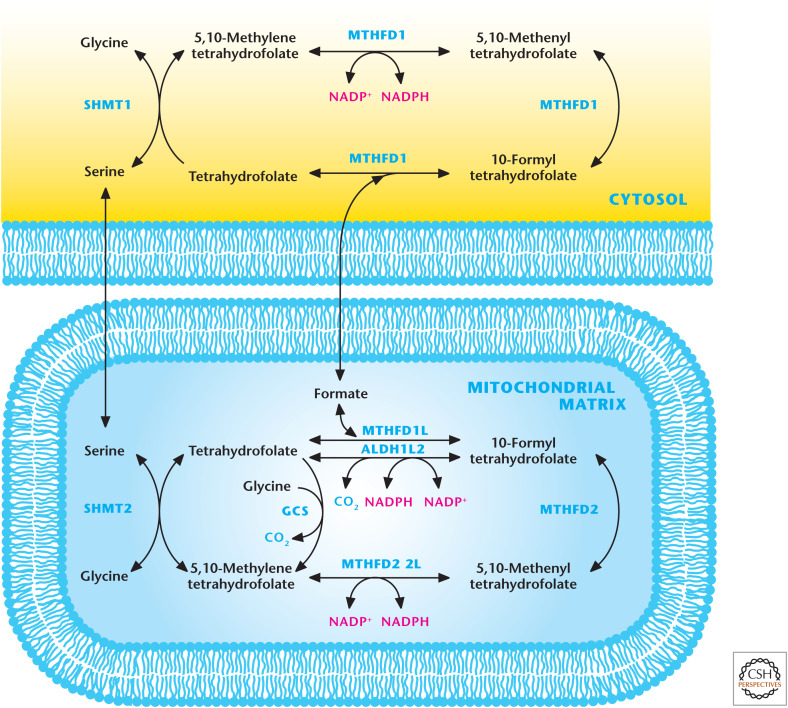
Mitochondrial and cytosolic one-carbon metabolism generates NADPH. The one-carbon amino acids serine and glycine can feed into folate cycle to generate NADPH. The cytosolic enzyme MTHFD1 and mitochondrial matrix enzymes MTHFD1L, MTHFD2L, and ALDH1L2 generate NADPH. One-carbon metabolism is a significant source of NADPH in the mitochondria. GCS, glycine cleavage system.
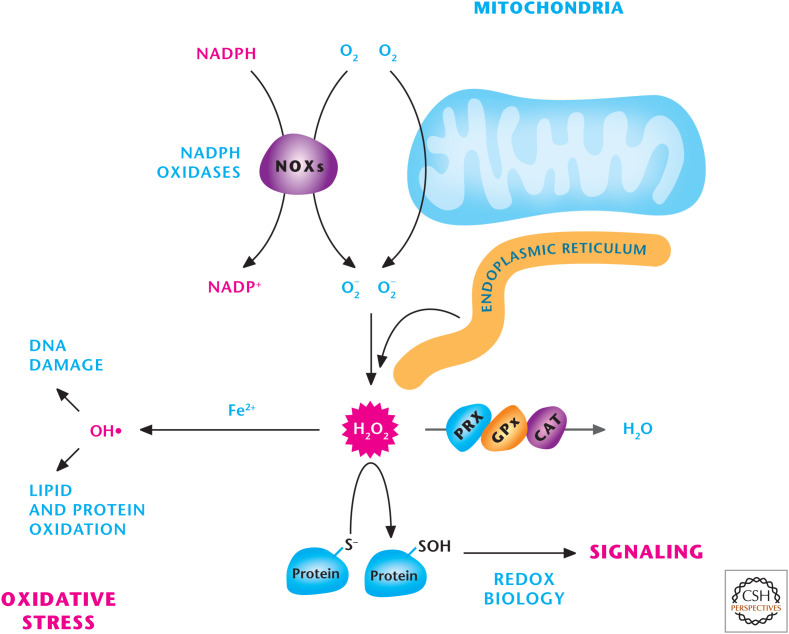
Sources of ROS. Mitochondria, NADPH oxidase (NOXs), and the endoplasmic reticulum are three major sources of H2O2, which can either be detoxified into H2O by peroxiredoxins (PRXs), catalase (CAT), or glutathione peroxidases (GPXs) or activate signaling pathways by oxidizing specific thiols within proteins. H2O2 in the presence of iron (Fe2+) can generate hydroxyl radical (OH−) that incurs damage to DNA, lipids, and proteins.
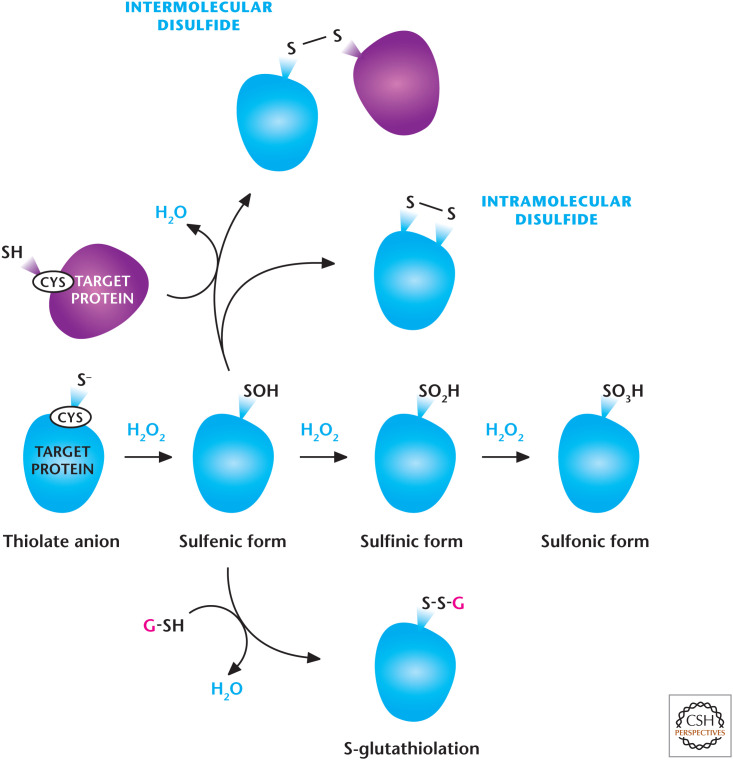
H2O2-dependent signaling. H2O2 regulates signaling pathways by oxidation of thiol groups on cysteines within proteins that show a low pKa, allowing the cysteine thiol group to exist as a thiolate anion (S−). H2O2 readily oxidizes thiolate, yielding SO−. Under high concentrations of H2O2, SO− can undergo further oxidation to generate SO2− and SO3−. SO− can undergo further modifications including intra- or intermolecular disulfide bonds and S-gluthathiolation. (Modified from Finkel 2011.)
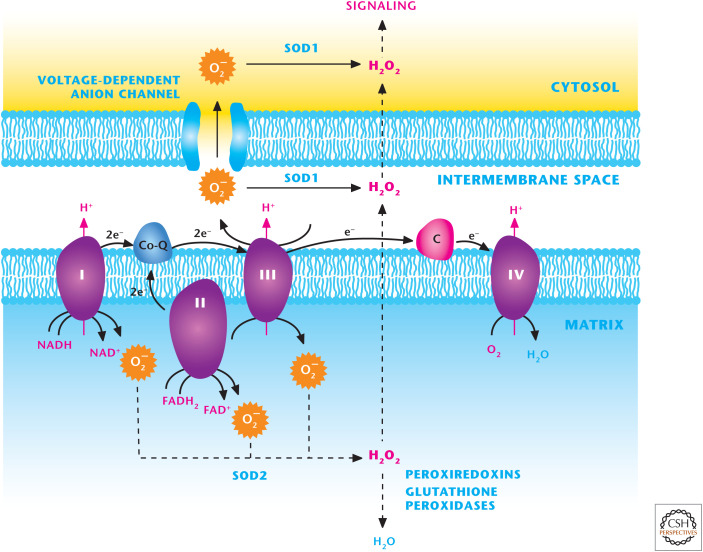
The mitochondrial electron transport chain (ETC) generates ROS. ETC complexes I, II, and III generate O2− superoxide in the mitochondrial matrix that is rapidly converted into H2O2 by SOD2. H2O2 can traverse through the inner and outer mitochondrial membranes to activate cellular signaling. Complex III can also generate O2− superoxide into the intermembrane space where it can traverse through voltage-dependent anion channels (VDACs) into the cytosol or into H2O2 by SOD1 in the intermembrane space. Subsequently H2O2 can efflux into the cytosol to activate signaling.
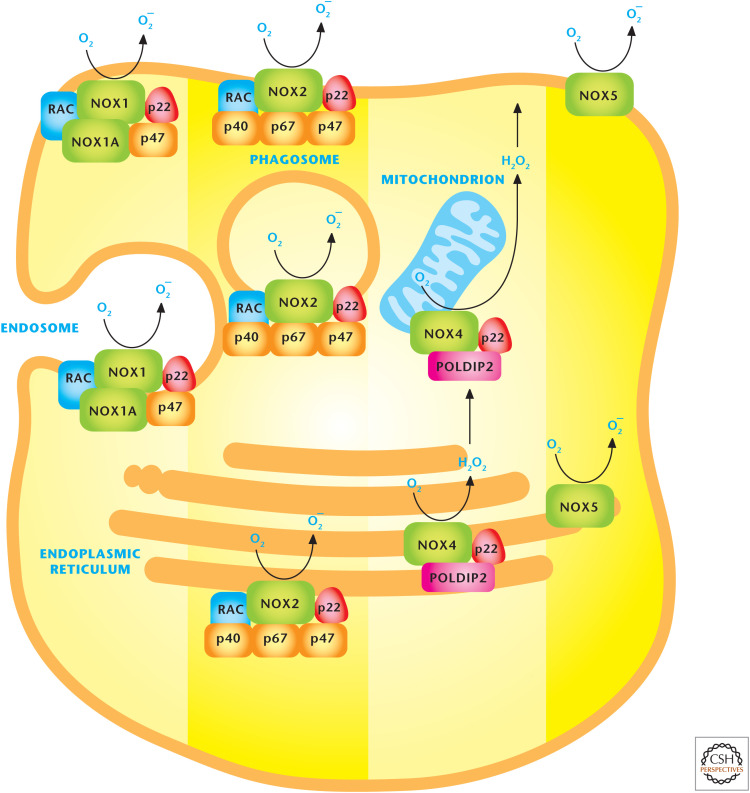
NADPH oxidases (NOXs) produce ROS. The NOX family of proteins localizes to the plasma, endoplasmic reticulum, and mitochondrial membranes. NOX catalytic subunits transfer electrons from NADPH across the membranes to molecular oxygen to generate O2− superoxide or H2O2 hydrogen peroxide. (Adapted, with permission, from Drummond et al. 2011, © Macmillan.)
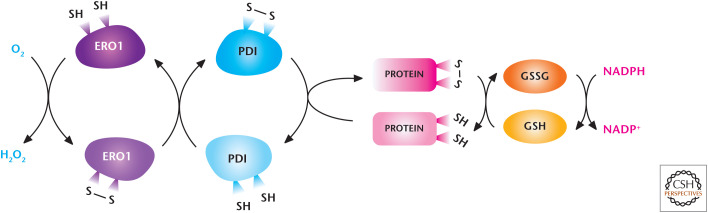
The endoplasmic reticulum (ER) generates ROS. The oxido-reductases PDI and ERO1 are coupled to generate H2O2 and mediate the disulfide bridge formation and protein folding of proteins. Reduced GSH reduces incorrectly placed disulfide bonds so that the protein can be recycled for correct disulfide bridge formation and folding. (Modified from Bhandary et al. 2013.)
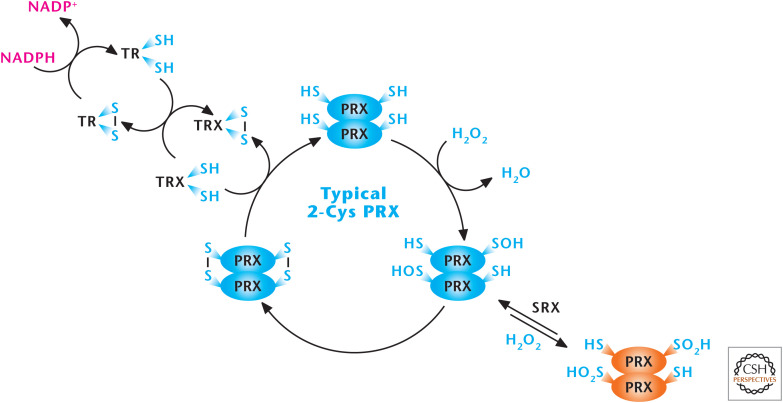
Peroxiredoxins (PRXs) scavenge H2O2. The peroxiredoxin (PRX1–6) family of proteins can be divided into three classes (typical 2-Cys, atypical 2-Cys [PRX5], and 1-Cys). PRXs function by undergoing oxidation by H2O2 at an active-site redox-sensitive cysteine and then subsequent reduction by cytosolic and mitochondrial thioredoxins (TRX1 and TRX2), cytosolic and mitochondrial thioredoxin reductase (TR1 and TR2), and NADPH. (Modified, with permission, from D'Autréaux and Toledano 2007, © Macmillan.)
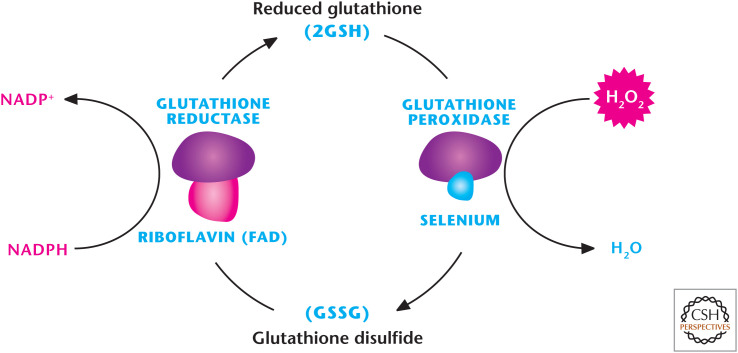
Glutathione peroxidase (GPX) detoxifies H2O2. There are eight mammalian GPXs known to catalyze the reduction of H2O2 to H2O by oxidizing GSH to GSSG. GSH reductase reduces GSSH back to GSH by using NADPH as an electron donor.
References
-
- Berg JM, Tymoczko JL, Stryer L. 2012. Biochemistry, 7th ed. WH Freeman, New York.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources