

Features and mechanisms of canonical and noncanonical genomic imprinting
- PMID: 34074696
- PMCID: PMC8168557
- DOI: 10.1101/gad.348422.121
Features and mechanisms of canonical and noncanonical genomic imprinting
Abstract
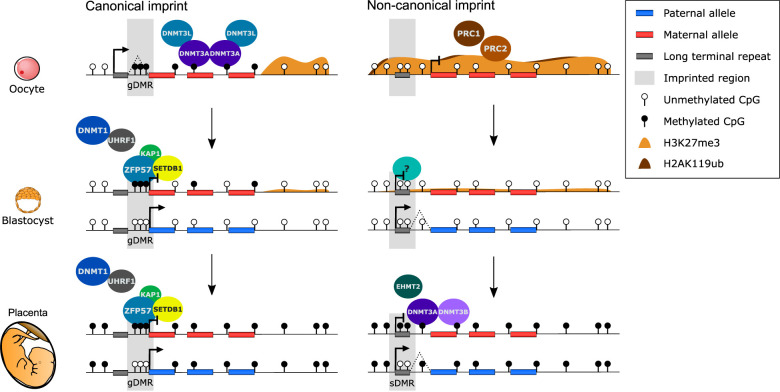
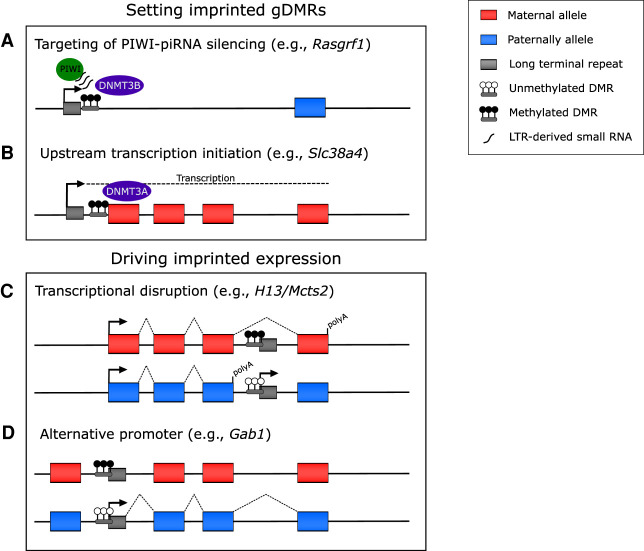
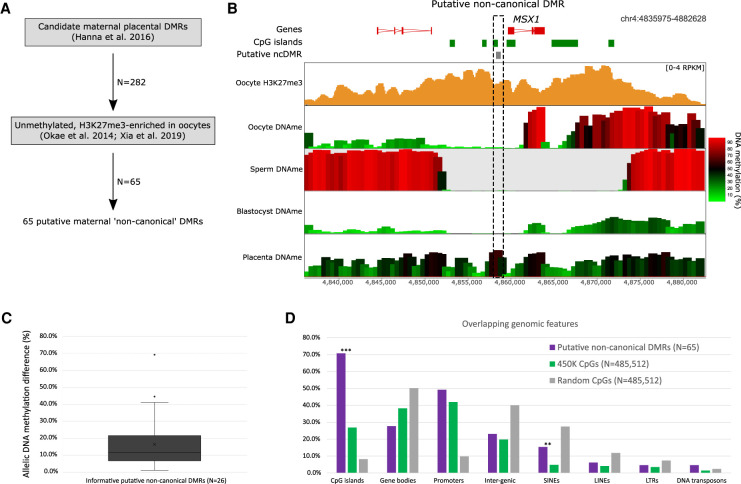
Genomic imprinting is the monoallelic expression of a gene based on parent of origin and is a consequence of differential epigenetic marking between the male and female germlines. Canonically, genomic imprinting is mediated by allelic DNA methylation. However, recently it has been shown that maternal H3K27me3 can result in DNA methylation-independent imprinting, termed "noncanonical imprinting." In this review, we compare and contrast what is currently known about the underlying mechanisms, the role of endogenous retroviral elements, and the conservation of canonical and noncanonical genomic imprinting.
Keywords: DNA methylation; chromatin; endogenous retroviruses; epigenetics; genomic imprinting.
© 2021 Hanna and Kelsey; Published by Cold Spring Harbor Laboratory Press.
Figures
References
-
- Anvar Z, Cammisa M, Riso V, Baglivo I, Kukreja H, Sparago A, Girardot M, Lad S, De Feis I, Cerrato F, et al. 2016. ZFP57 recognizes multiple and closely spaced sequence motif variants to maintain repressive epigenetic marks in mouse embryonic stem cells. Nucleic Acids Res 44: 1118–1132. 10.1093/nar/gkv1059 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources