

Reliance on Cox10 and oxidative metabolism for antigen-specific NK cell expansion
- PMID: 34077722
- PMCID: PMC8229496
- DOI: 10.1016/j.celrep.2021.109209
Reliance on Cox10 and oxidative metabolism for antigen-specific NK cell expansion
Abstract
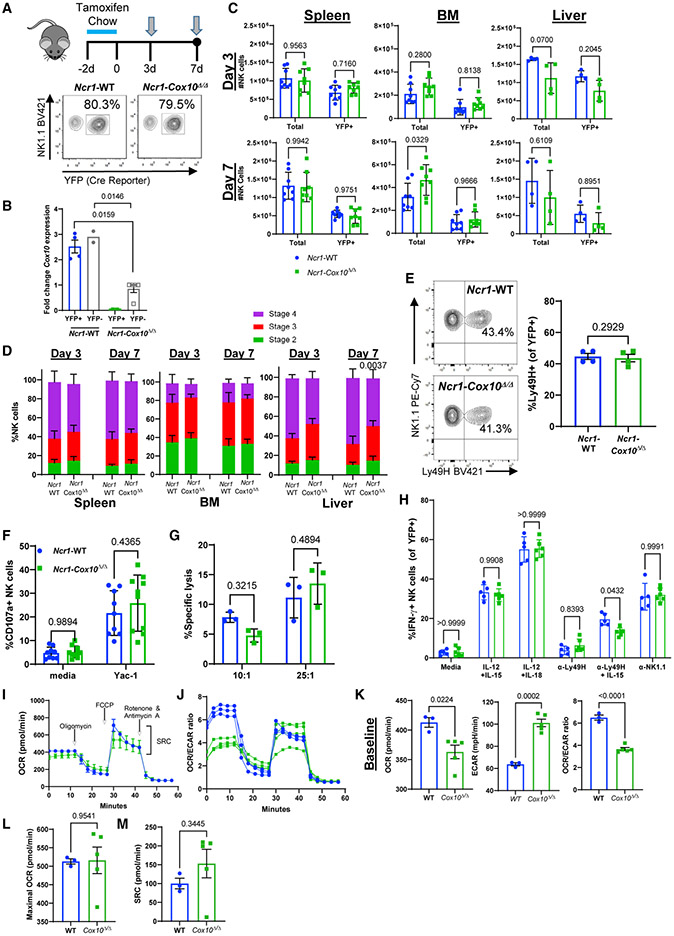
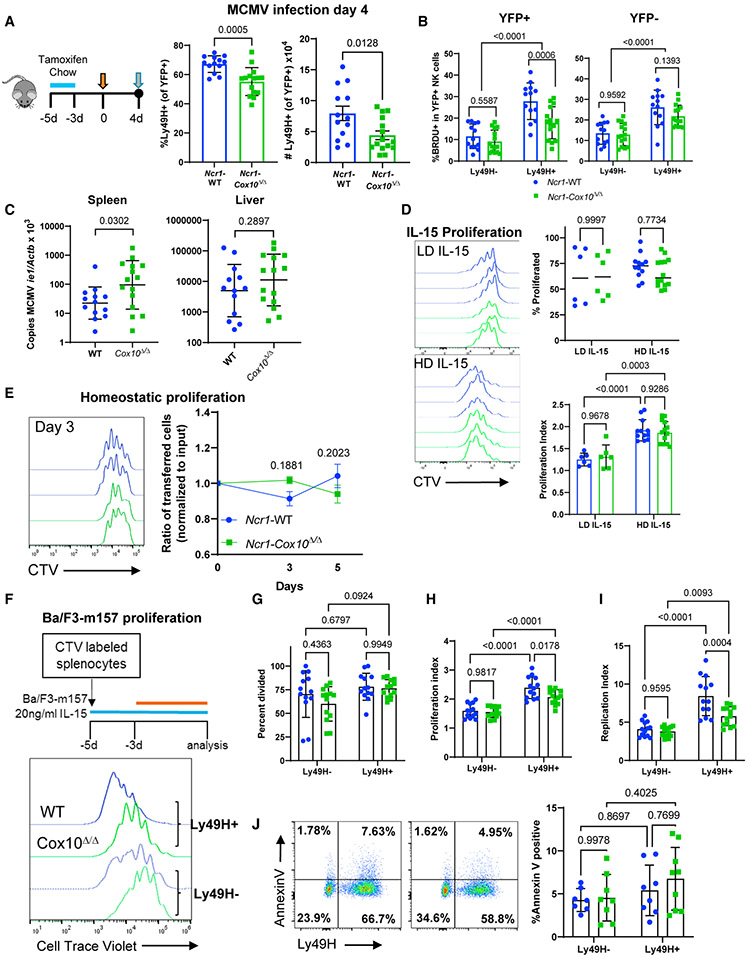
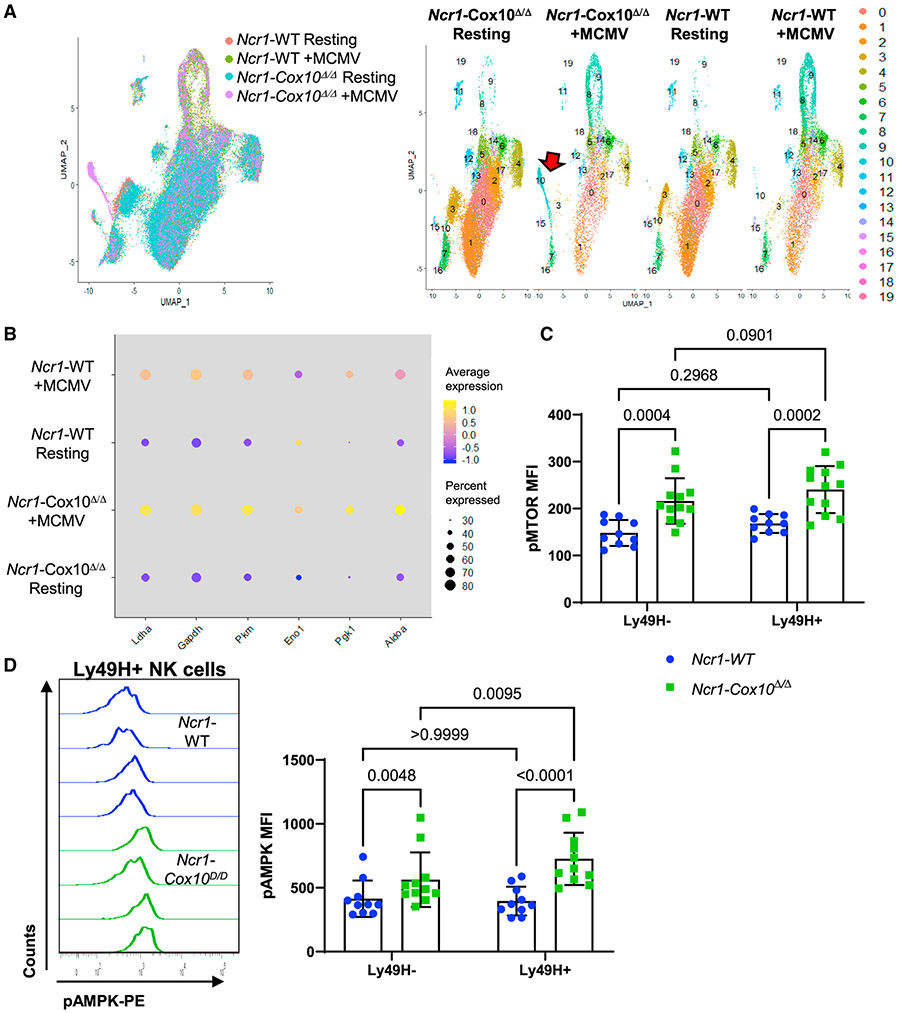
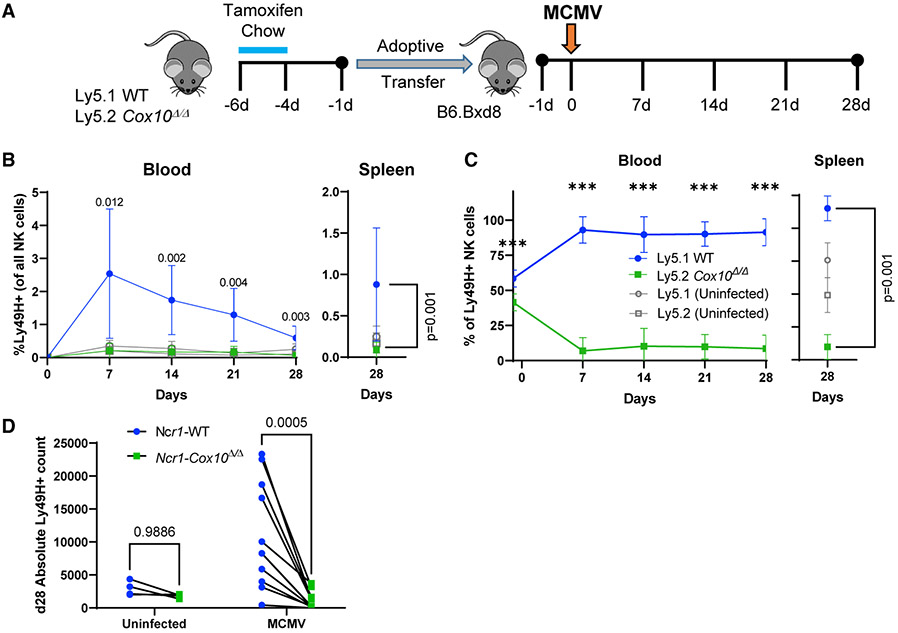
Natural killer (NK) cell effector functions are dependent on metabolic regulation of cellular function; however, less is known about in vivo metabolic pathways required for NK cell antiviral function. Mice with an inducible NK-specific deletion of Cox10, which encodes a component of electron transport chain complex IV, were generated to investigate the role of oxidative phosphorylation in NK cells during murine cytomegalovirus (MCMV) infection. Ncr1-Cox10Δ/Δ mice had normal numbers of NK cells but impaired expansion of antigen-specific Ly49H+ NK cells and impaired NK cell memory formation. Proliferation in vitro and homeostatic expansion were intact, indicating a specific metabolic requirement for antigen-driven proliferation. Cox10-deficient NK cells upregulated glycolysis, associated with increased AMP-activated protein kinase (AMPK) and mammalian target of rapamycin (mTOR) activation, although this was insufficient to protect the host. These data demonstrate that oxidative metabolism is required for NK cell antiviral responses in vivo.
Keywords: Cox10; NK cells; metabolism; murine cytomegalovirus; oxidative phosphorylation; proliferation.
Copyright © 2021 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Adams NM, Grassmann S, and Sun JC (2020). Clonal expansion of innate and adaptive lymphocytes. Nat. Rev. Immunol 20, 694–707. - PubMed
-
- Antonicka H, Leary SC, Guercin GH, Agar JN, Horvath R, Kennaway NG, Harding CO, Jaksch M, and Shoubridge EA (2003). Mutations in COX10 result in a defect in mitochondrial heme A biosynthesis and account for multiple, early-onset clinical phenotypes associated with isolated COX deficiency. Hum. Mol. Genet 12, 2693–2702. - PubMed
-
- Arase H, Mocarski ES, Campbell AE, Hill AB, and Lanier LL (2002). Direct recognition of cytomegalovirus by activating and inhibitory NK cell receptors. Science 296, 1323–1326. - PubMed
-
- Assmann N, O’Brien KL, Donnelly RP, Dyck L, Zaiatz-Bittencourt V, Loftus RM, Heinrich P, Oefner PJ, Lynch L, Gardiner CM, et al. (2017). Srebp-controlled glucose metabolism is essential for NK cell functional responses. Nat. Immunol 18, 1197–1206. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
