

Protective neutralizing antibodies from human survivors of Crimean-Congo hemorrhagic fever
- PMID: 34077751
- PMCID: PMC8559771
- DOI: 10.1016/j.cell.2021.05.001
Protective neutralizing antibodies from human survivors of Crimean-Congo hemorrhagic fever
Abstract
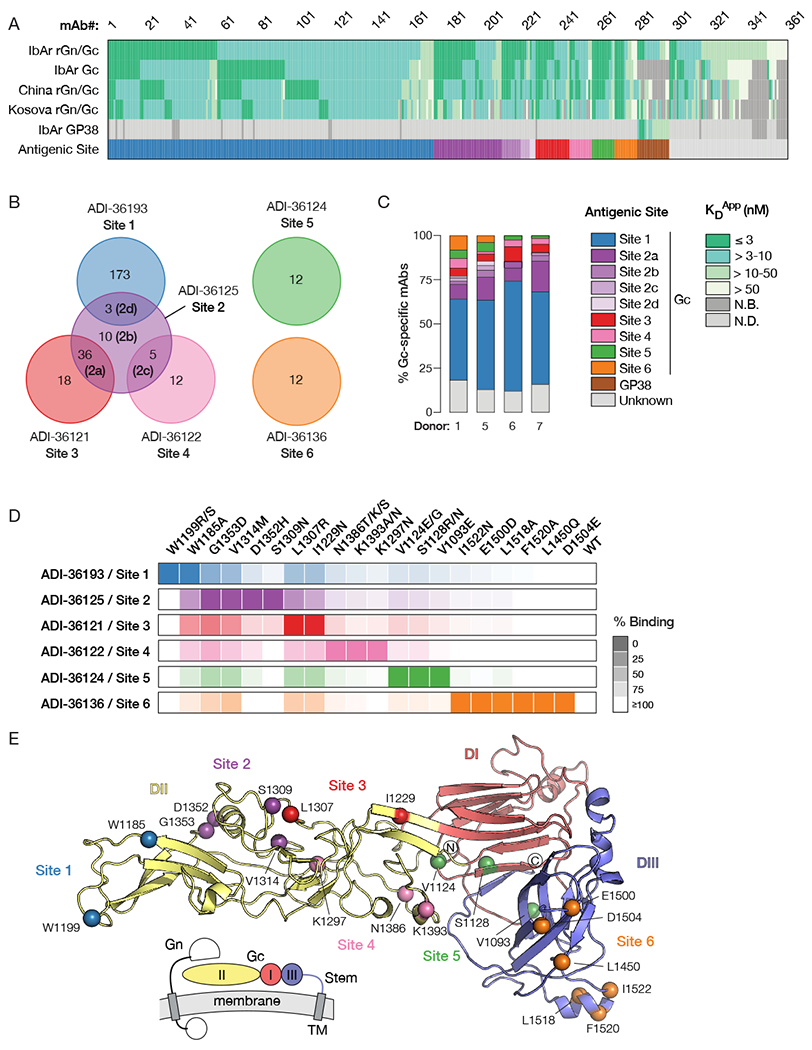
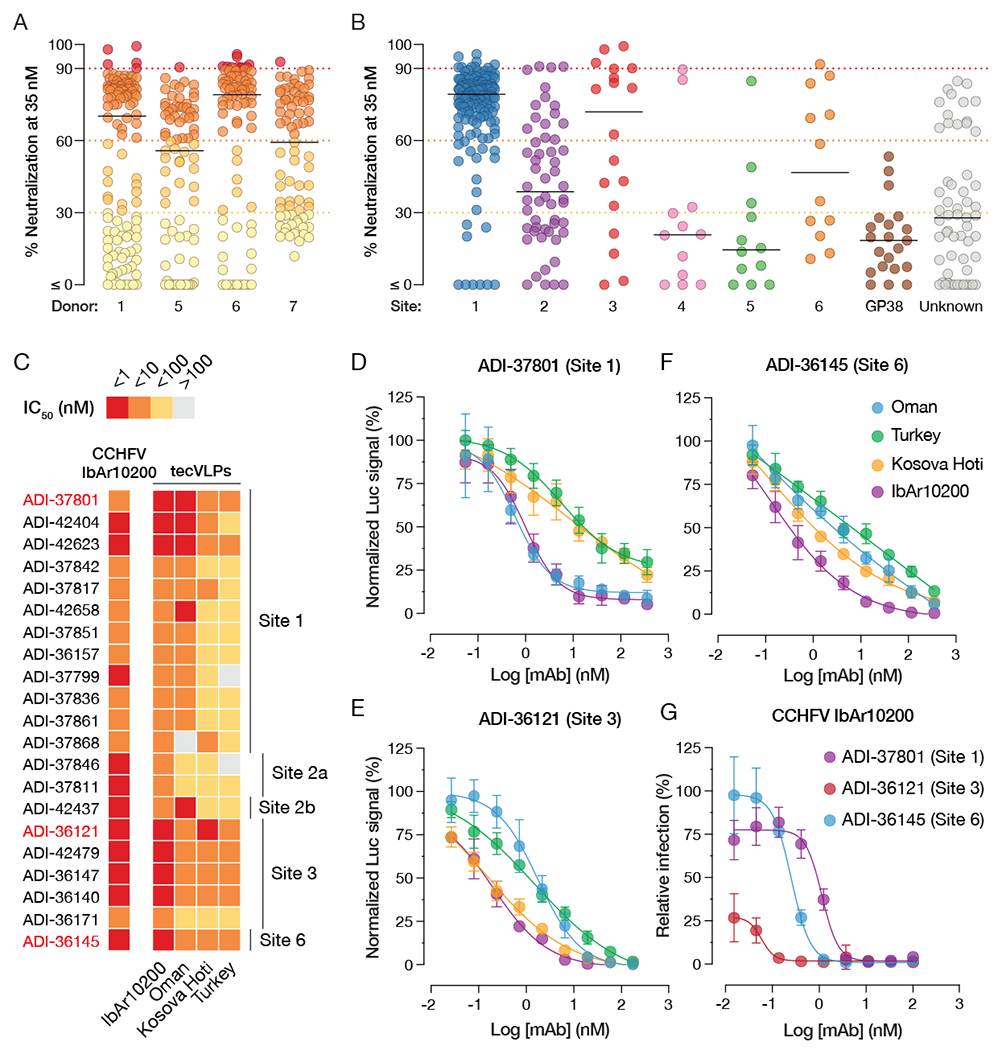
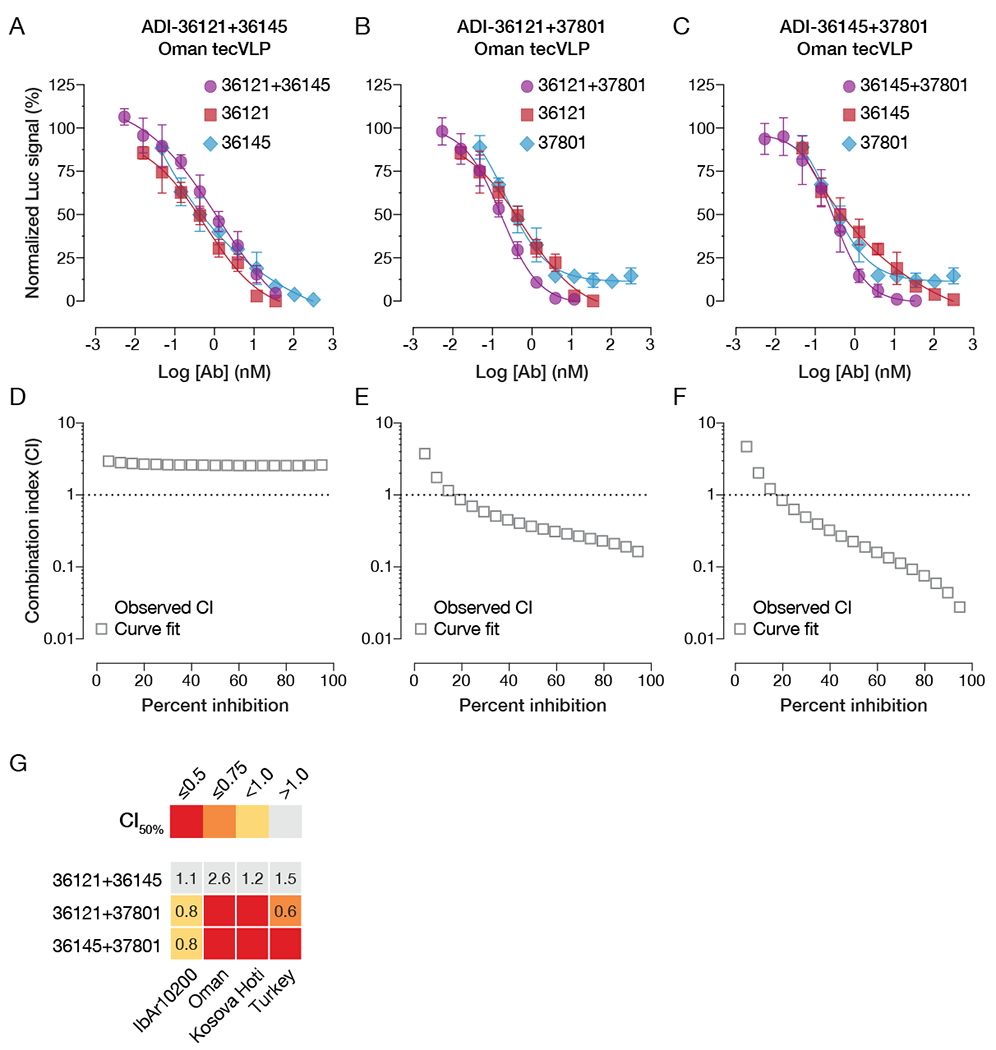
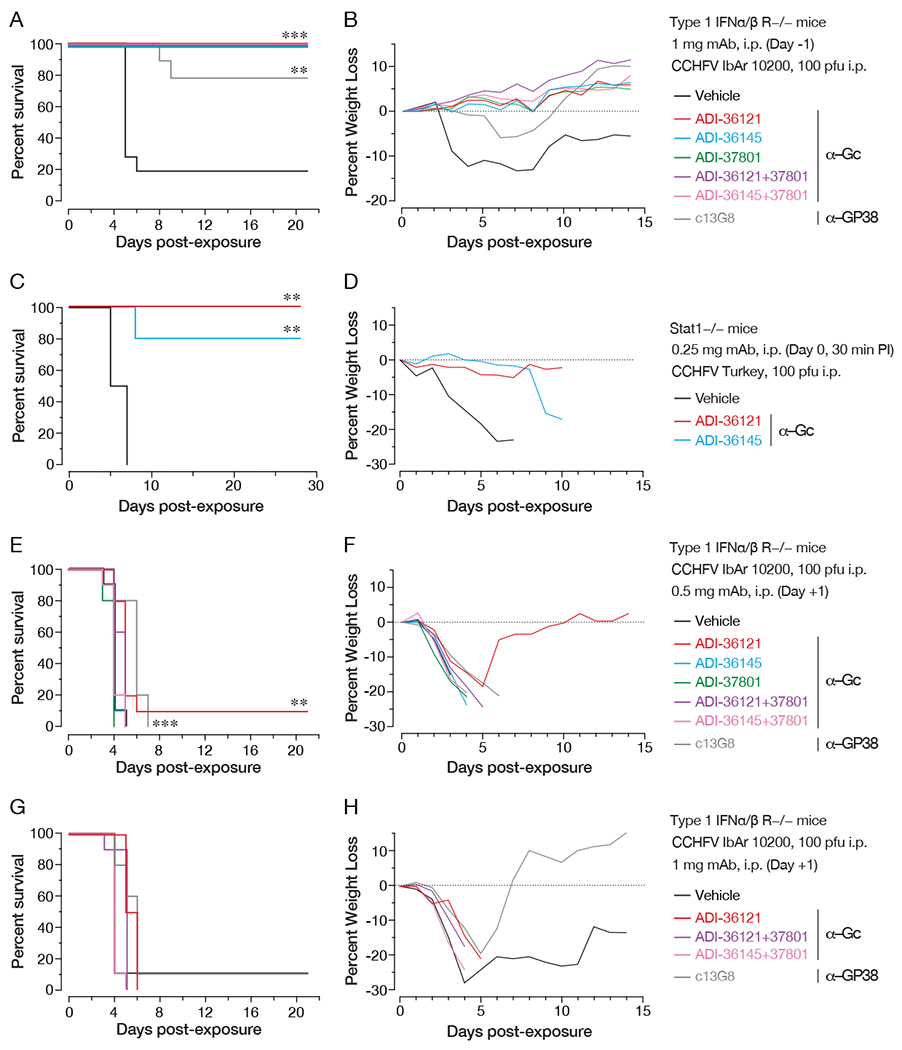
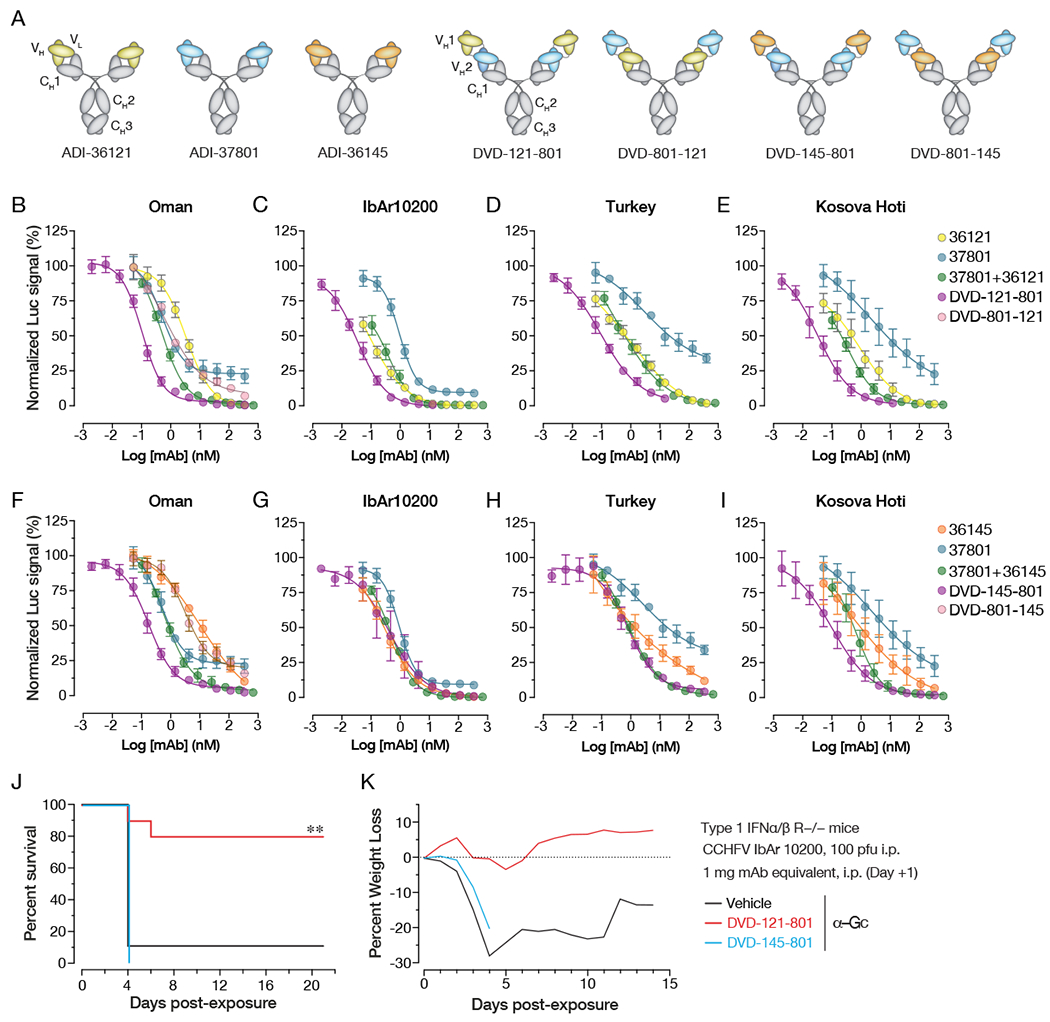
Crimean-Congo hemorrhagic fever virus (CCHFV) is a World Health Organization priority pathogen. CCHFV infections cause a highly lethal hemorrhagic fever for which specific treatments and vaccines are urgently needed. Here, we characterize the human immune response to natural CCHFV infection to identify potent neutralizing monoclonal antibodies (nAbs) targeting the viral glycoprotein. Competition experiments showed that these nAbs bind six distinct antigenic sites in the Gc subunit. These sites were further delineated through mutagenesis and mapped onto a prefusion model of Gc. Pairwise screening identified combinations of non-competing nAbs that afford synergistic neutralization. Further enhancements in neutralization breadth and potency were attained by physically linking variable domains of synergistic nAb pairs through bispecific antibody (bsAb) engineering. Although multiple nAbs protected mice from lethal CCHFV challenge in pre- or post-exposure prophylactic settings, only a single bsAb, DVD-121-801, afforded therapeutic protection. DVD-121-801 is a promising candidate suitable for clinical development as a CCHFV therapeutic.
Keywords: CCHFV; Crimean-Congo hemorrhagic fever virus; antibody therapeutic; bunyavirus; emerging virus; human monoclonal antibody; monoclonal antibody; nairovirus; tickborne; viral glycoprotein.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests K.C. is a scientific advisory board member of Integrum Scientific and Biovaxys Technology Corporation. K.C., J.S.M., and J.R.L. are scientific advisory board members of the Pandemic Security Initiative of Celdara Medical. N.T.P. and L.M.W. are employees and shareholders of Adimab. D.P.M. is a shareholder of Adimab. Z.A.B., D.M.A., C.L.M., and L.Z. are shareholders and employees of Mapp Biopharmaceutical. Mapp Biopharmaceutical has filed a patent application related to this work.
Figures
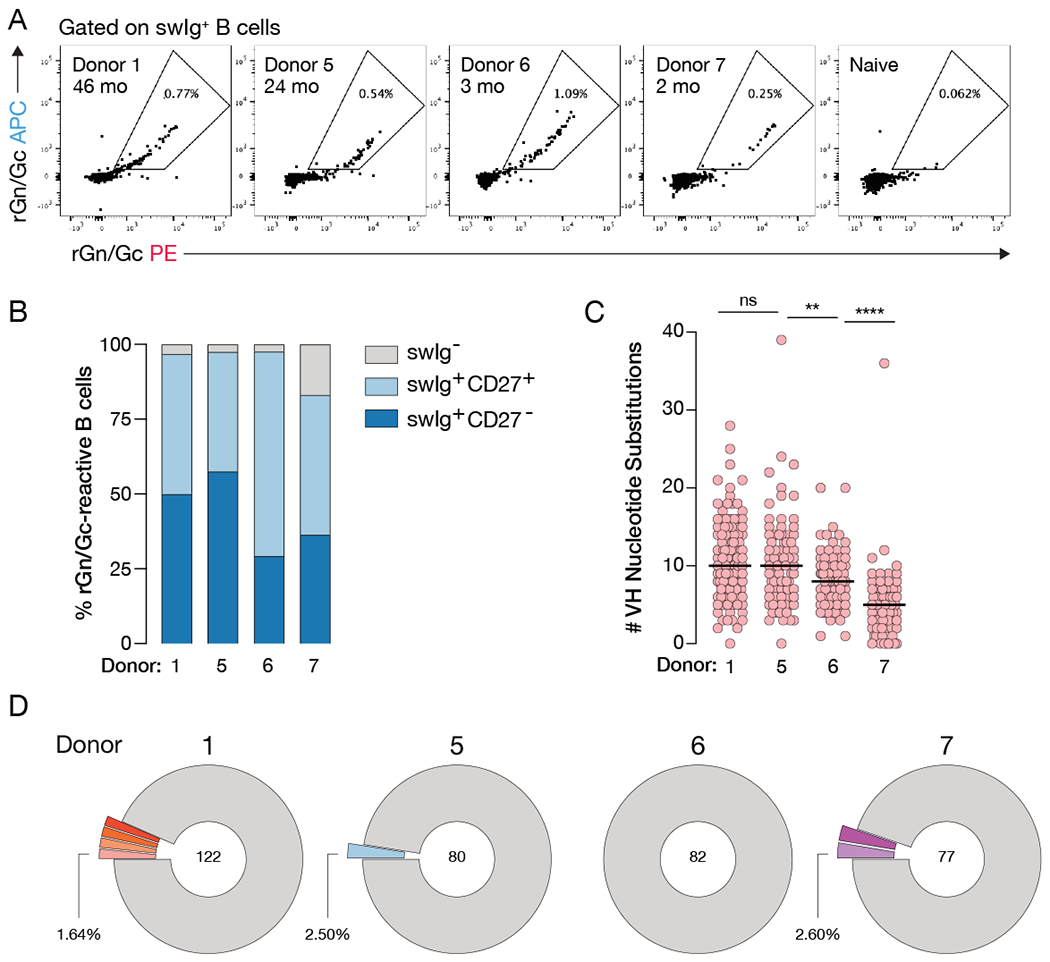
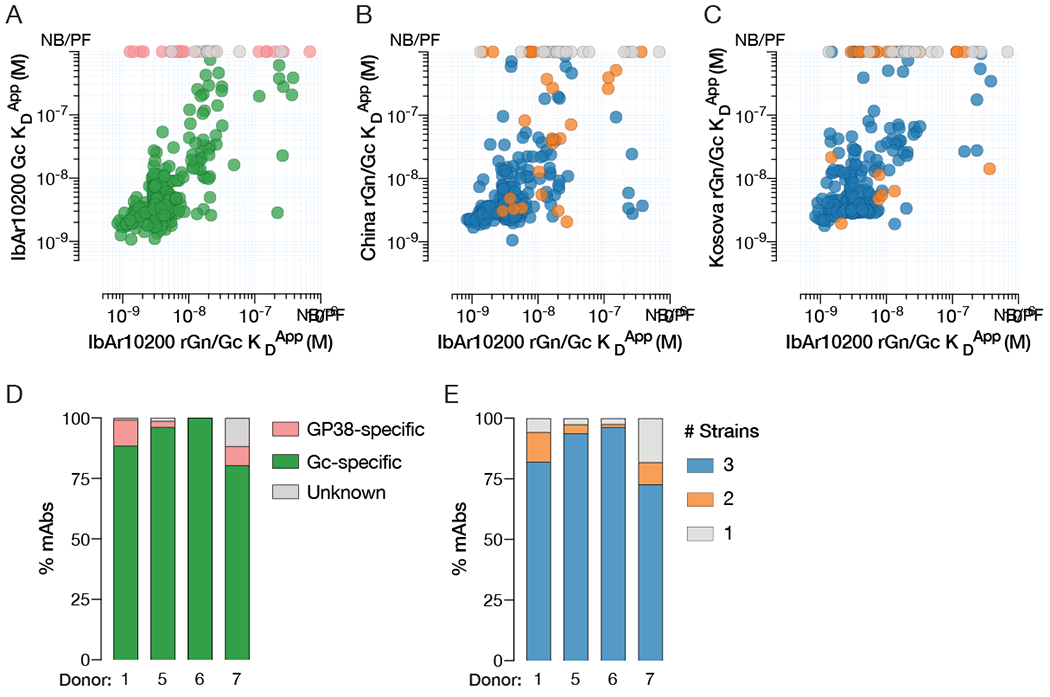
References
-
- Andrews SF, Chambers MJ, Schramm CA, Plyler J, Raab JE, Kanekiyo M, Gillespie RA, Ransier A, Darko S, Hu J, et al. (2019). Activation Dynamics and Immunoglobulin Evolution of Pre-existing and Newly Generated Human Memory B cell Responses to Influenza Hemagglutinin. Immunity 51, 398–410.e5. - PubMed
-
- Beltramello M, Williams KL, Simmons CP, Macagno A, Simonelli L, Quyen NTH, Sukupolvi-Petty S, Navarro-Sanchez E, Young PR, de Silva AM, et al. (2010). The human immune response to Dengue virus is dominated by highly cross-reactive antibodies endowed with neutralizing and enhancing activity. Cell Host Microbe 8, 271–283. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
