

Live imaging of chromatin distribution reveals novel principles of nuclear architecture and chromatin compartmentalization
- PMID: 34078602
- PMCID: PMC8172175
- DOI: 10.1126/sciadv.abf6251
Live imaging of chromatin distribution reveals novel principles of nuclear architecture and chromatin compartmentalization
Abstract
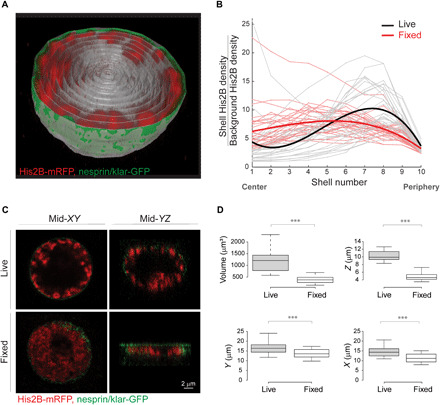
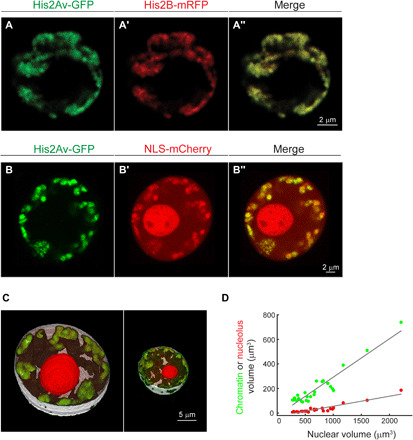
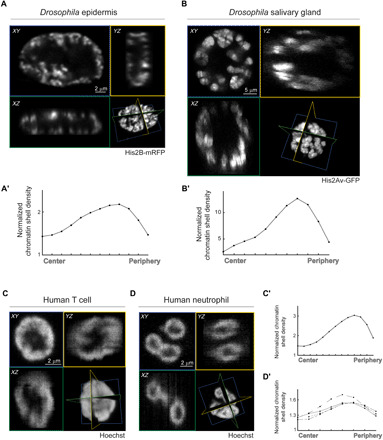
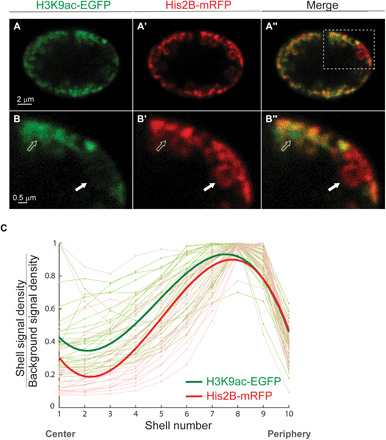
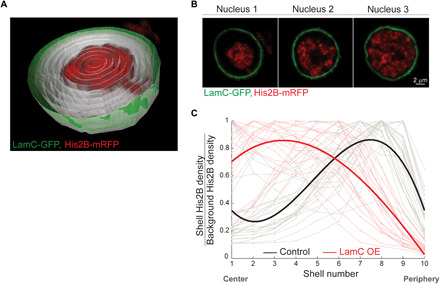
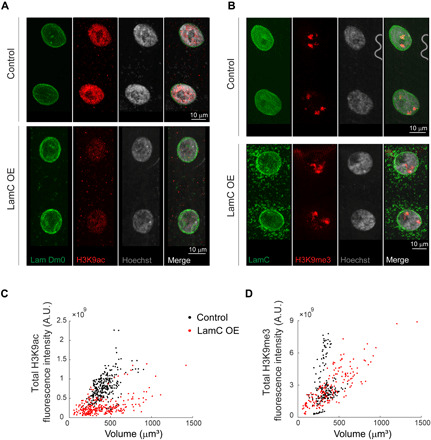
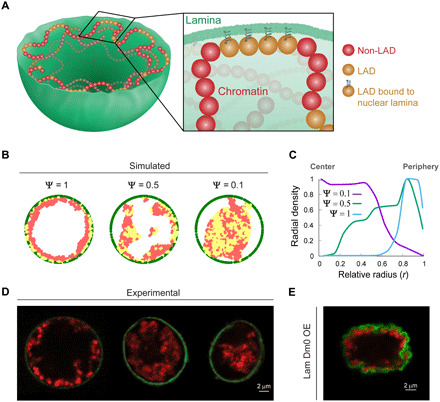
The three-dimensional organization of chromatin contributes to transcriptional control, but information about native chromatin distribution is limited. Imaging chromatin in live Drosophila larvae, with preserved nuclear volume, revealed that active and repressed chromatin separates from the nuclear interior and forms a peripheral layer underneath the nuclear lamina. This is in contrast to the current view that chromatin distributes throughout the nucleus. Furthermore, peripheral chromatin organization was observed in distinct Drosophila tissues, as well as in live human effector T lymphocytes and neutrophils. Lamin A/C up-regulation resulted in chromatin collapse toward the nuclear center and correlated with a significant reduction in the levels of active chromatin. Physical modeling suggests that binding of lamina-associated domains combined with chromatin self-attractive interactions recapitulate the experimental chromatin distribution profiles. Together, our findings reveal a novel mode of mesoscale organization of peripheral chromatin sensitive to lamina composition, which is evolutionary conserved.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution NonCommercial License 4.0 (CC BY-NC).
Figures
References
-
- Lakadamyali M., Cosma M. P., Visualizing the genome in high resolution challenges our textbook understanding. Nat. Methods 17, 371–379 (2020). - PubMed
-
- Ricci M. A., Manzo C., García-Parajo M. F., Lakadamyali M., Cosma M. P., Chromatin fibers are formed by heterogeneous groups of nucleosomes in vivo. Cell 160, 1145–1158 (2015). - PubMed
-
- Cremer T., Cremer C., Chromosome territories, nuclear architecture and gene regulation in mammalian cells. Nat. Rev. Genet. 2, 292–301 (2001). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
