

First exposure to the pandemic H1N1 virus induced broadly neutralizing antibodies targeting hemagglutinin head epitopes
- PMID: 34078743
- PMCID: PMC10173203
- DOI: 10.1126/scitranslmed.abg4535
First exposure to the pandemic H1N1 virus induced broadly neutralizing antibodies targeting hemagglutinin head epitopes
Erratum in
-
Erratum for the Research Article: "First exposure to the pandemic H1N1 virus induced broadly neutralizing antibodies targeting hemagglutinin head epitopes" by J. J. Guthmiller, J. Han, L. Li, A. W. Freyn, S. T. H. Liu, O. Stovicek, C. T. Stamper, H. L. Dugan, M. E. Tepora, H. A. Utset, D. J. Bitar, N. J. Hamel, S. Changrob, N.-Y. Zheng, M. Huang, F. Krammer, R. Nachbagauer, P. Palese, A. B. Ward, P. C. Wilson.Sci Transl Med. 2022 Feb 23;14(633):eabo6315. doi: 10.1126/scitranslmed.abo6315. Epub 2022 Feb 23. Sci Transl Med. 2022. PMID: 35196025 No abstract available.
Abstract
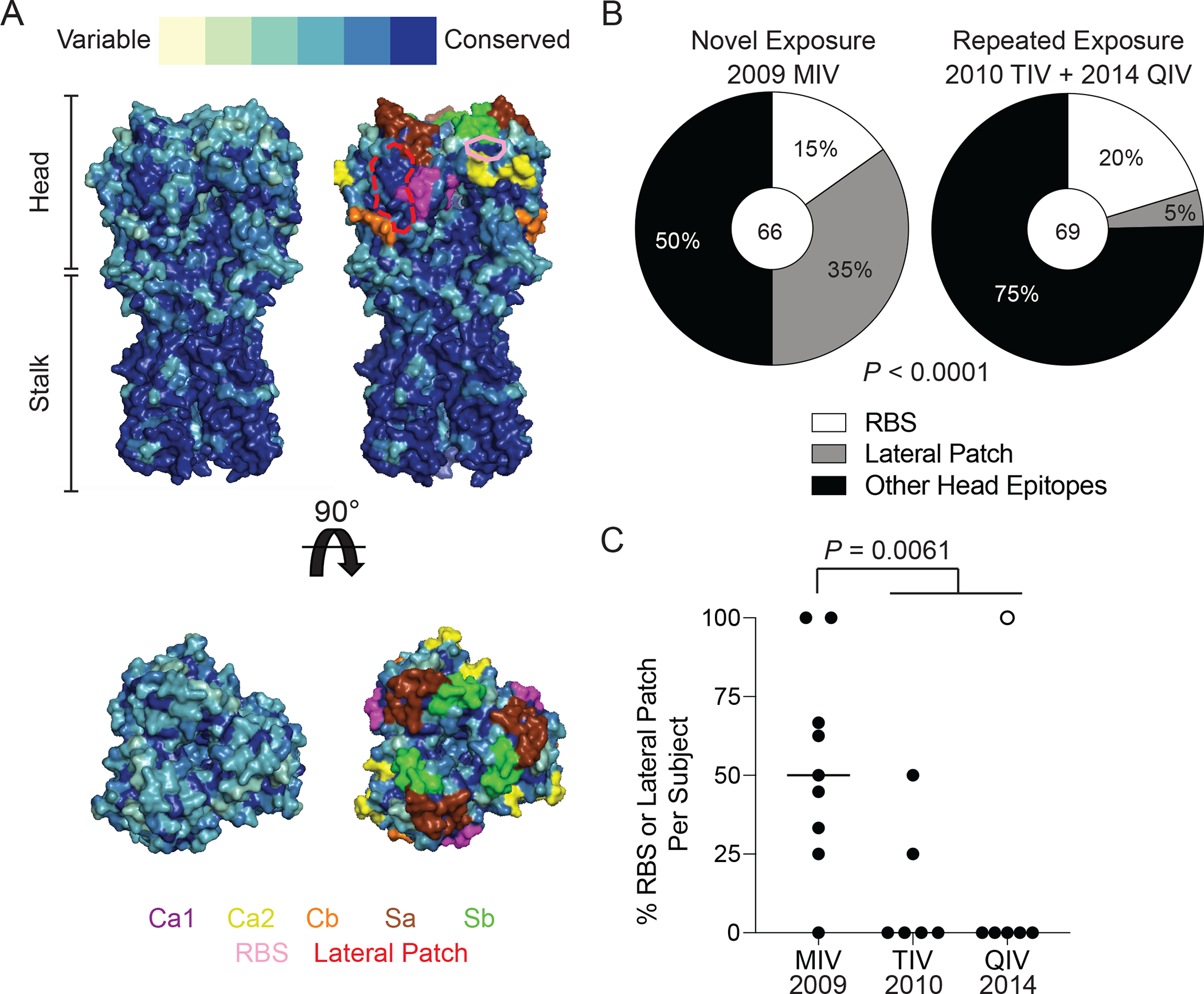
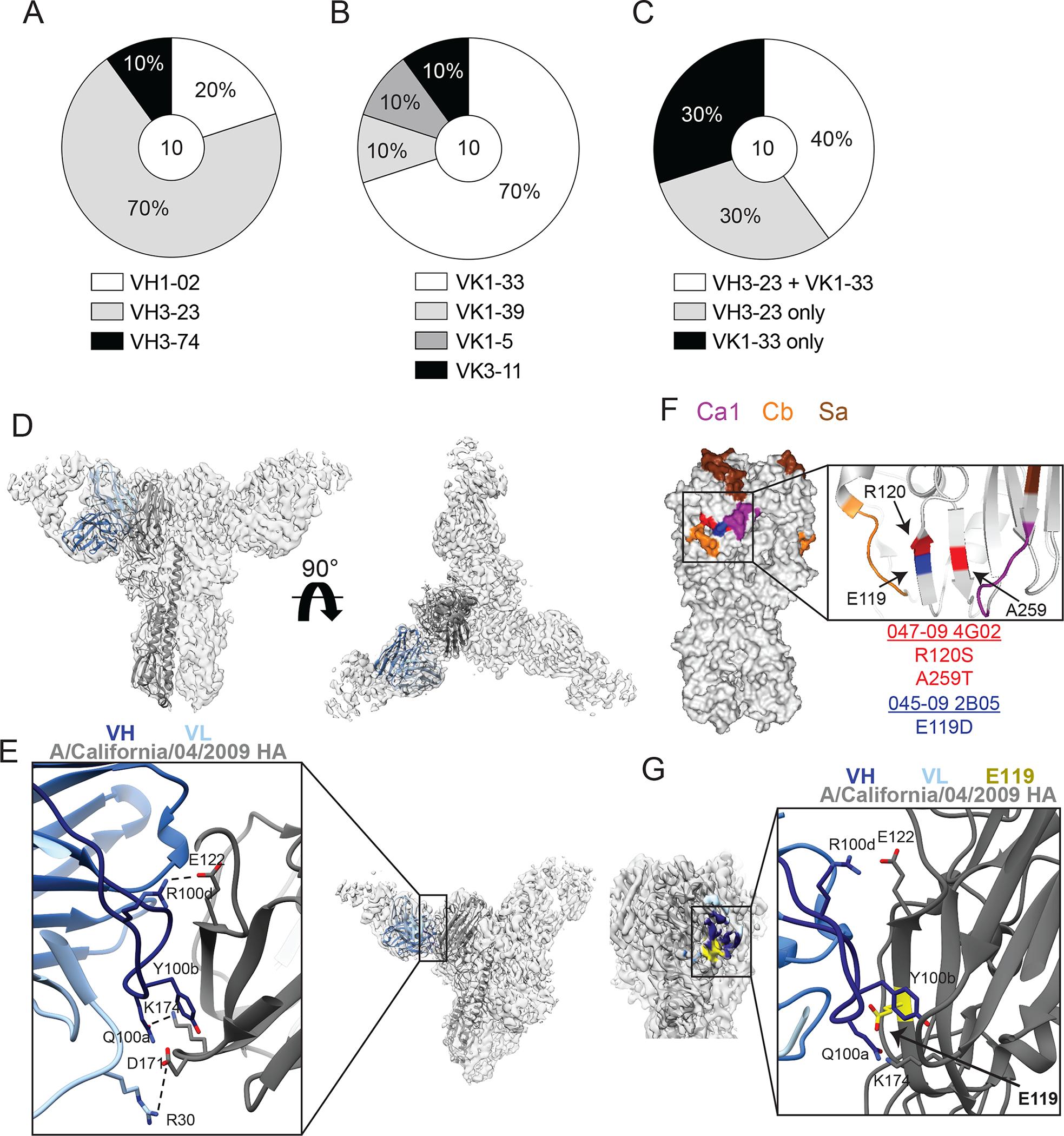
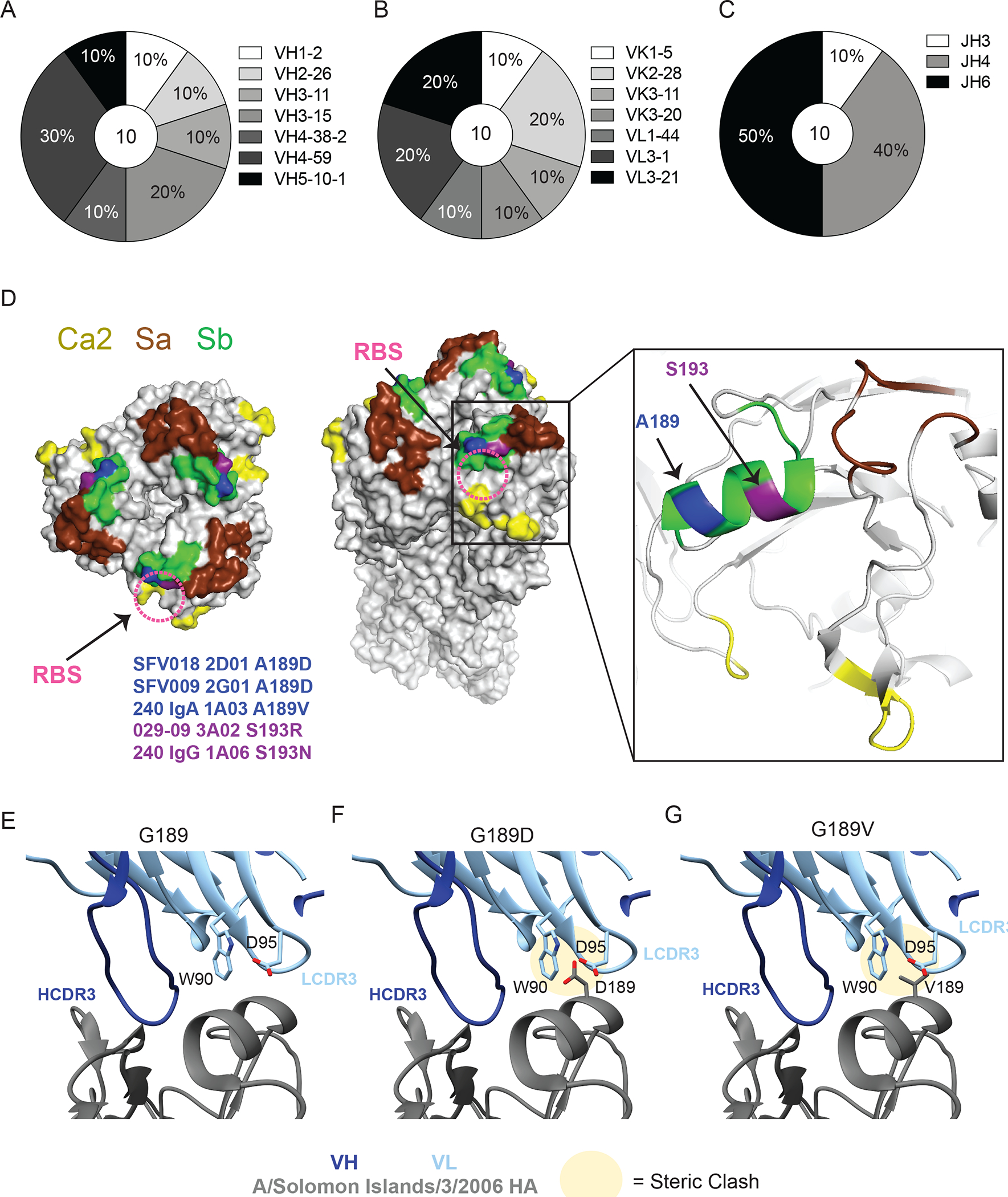
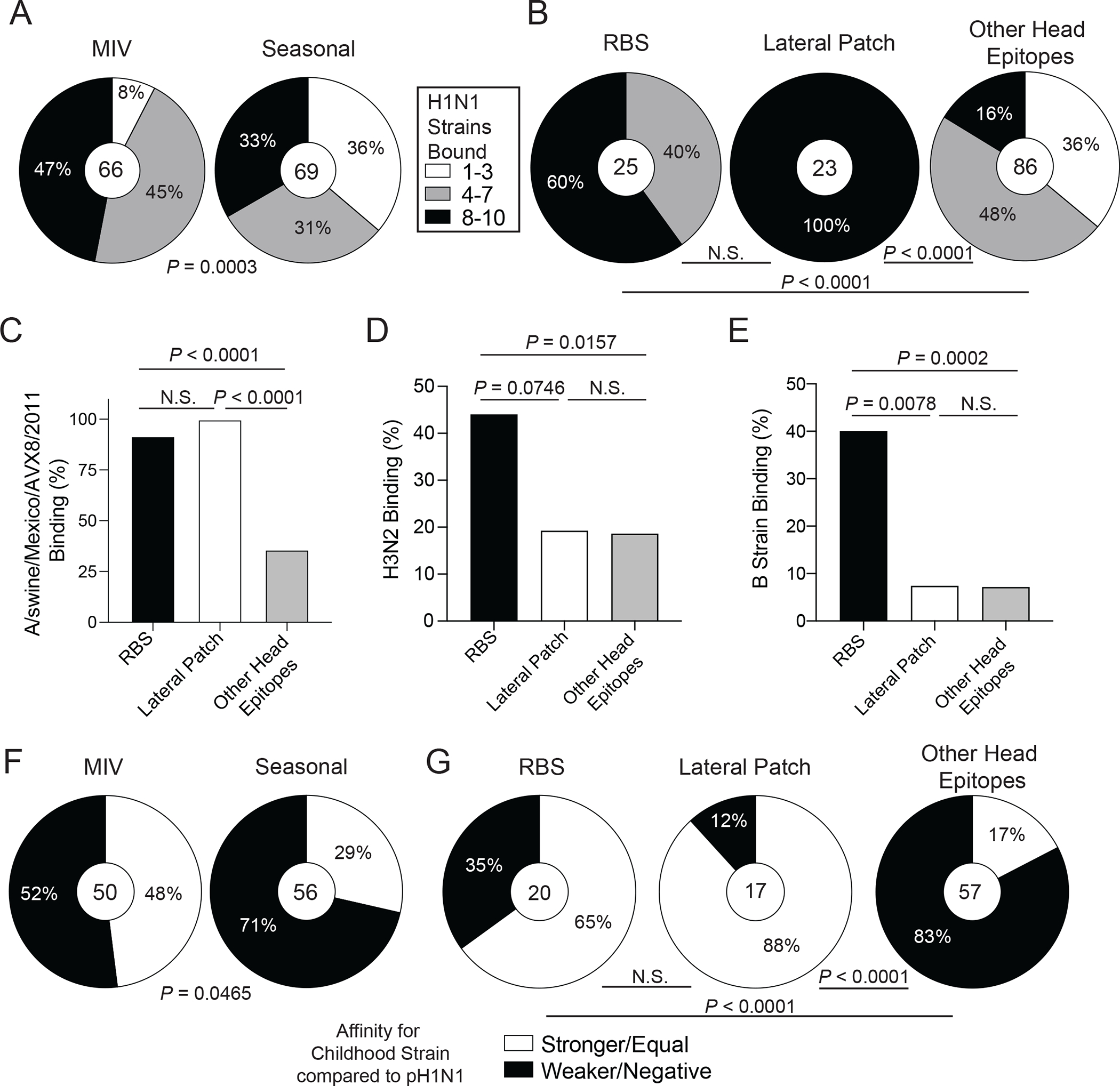
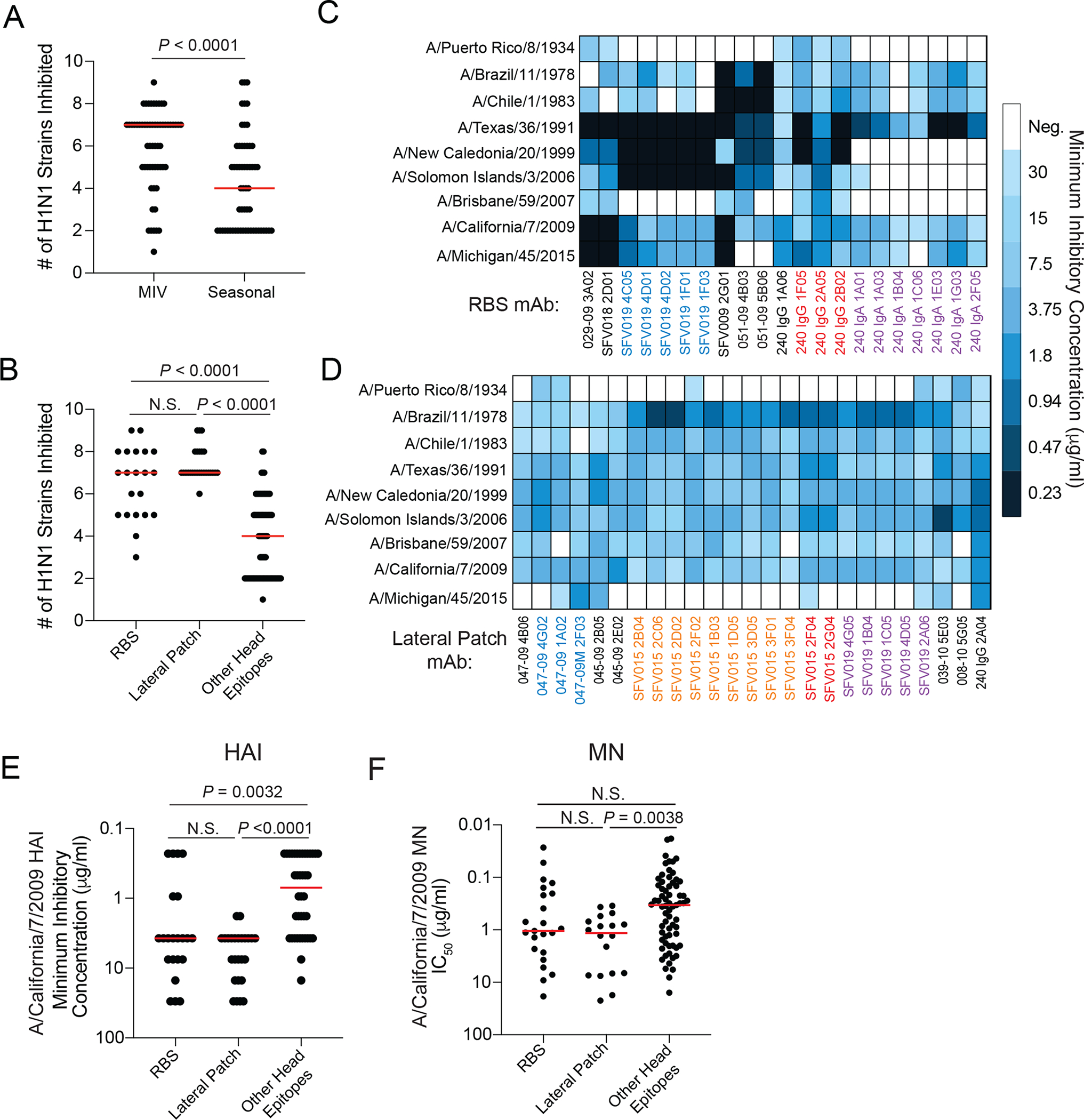
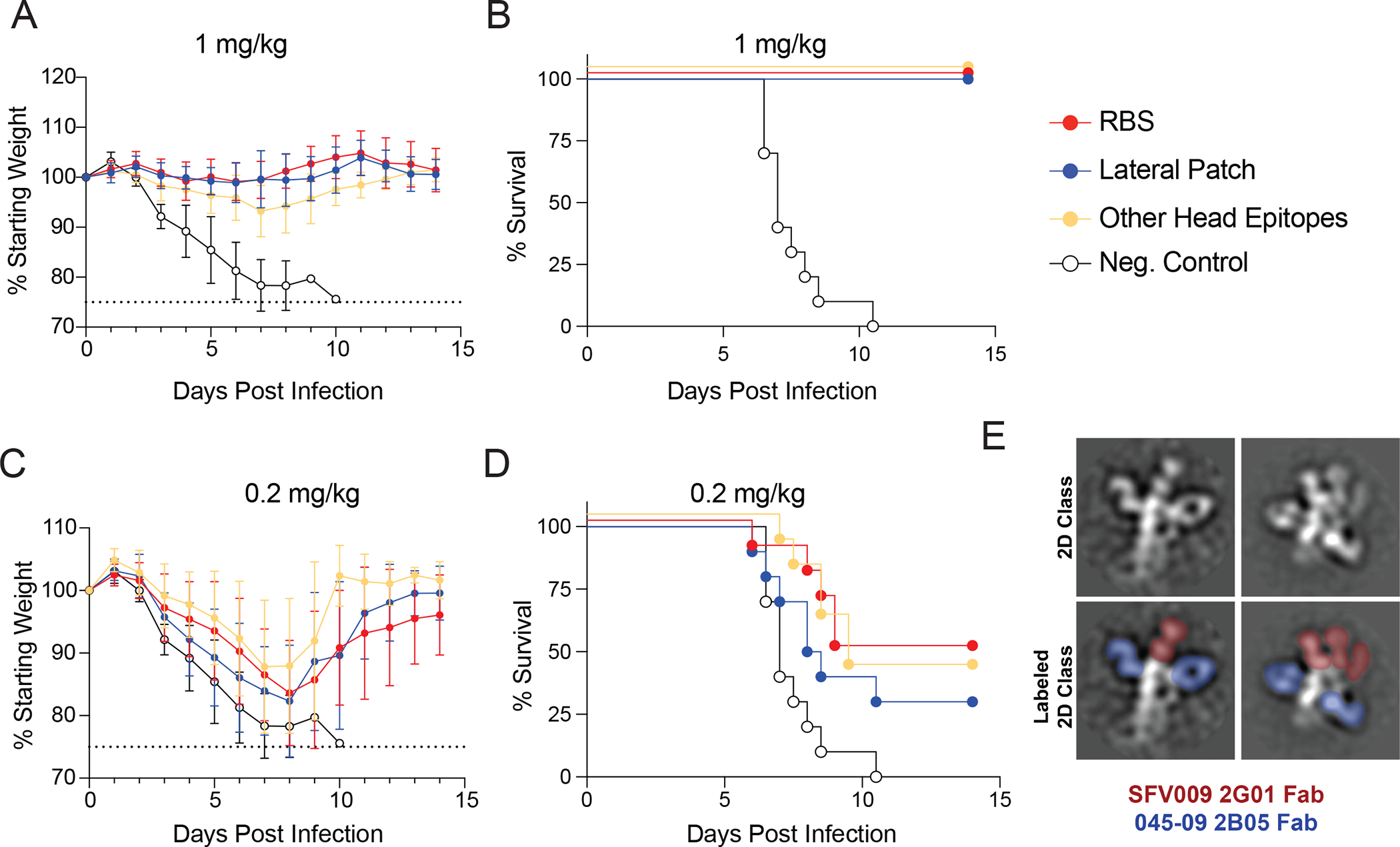
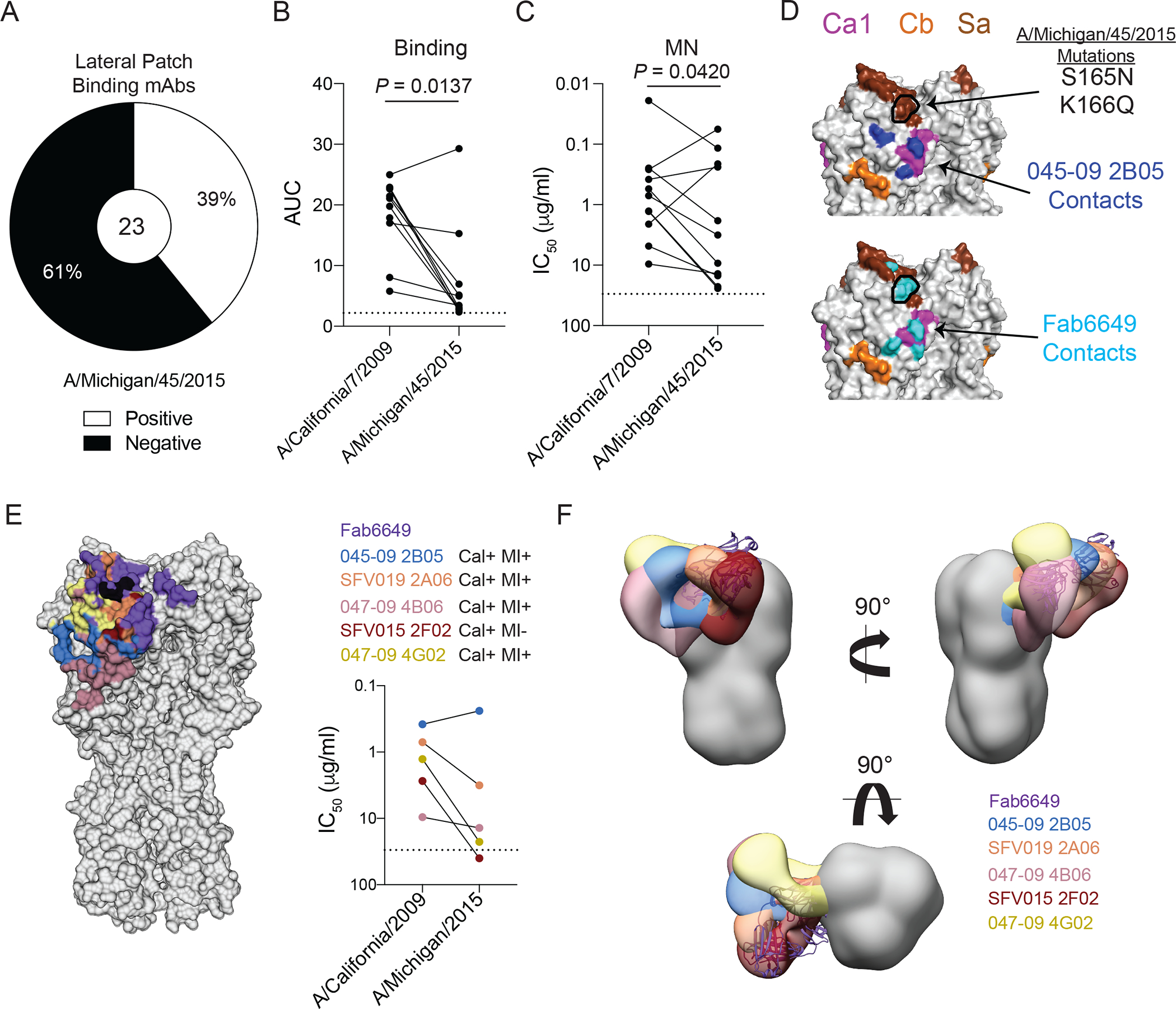
Broadly neutralizing antibodies are critical for protection against both drifted and shifted influenza viruses. Here, we reveal that first exposure to the 2009 pandemic H1N1 influenza virus recalls memory B cells that are specific to the conserved receptor-binding site (RBS) or lateral patch epitopes of the hemagglutinin (HA) head domain. Monoclonal antibodies (mAbs) generated against these epitopes are broadly neutralizing against H1N1 viruses spanning 40 years of viral evolution and provide potent protection in vivo. Lateral patch-targeting antibodies demonstrated near universal binding to H1 viruses, and RBS-binding antibodies commonly cross-reacted with H3N2 viruses and influenza B viruses. Lateral patch-targeting mAbs were restricted to expressing the variable heavy-chain gene VH3-23 with or without the variable kappa-chain gene VK1-33 and often had a Y-x-R motif within the heavy-chain complementarity determining region 3 to make key contacts with HA. Moreover, lateral patch antibodies that used both VH3-23 and VK1-33 maintained neutralizing capability with recent pH1N1 strains that acquired mutations near the lateral patch. RBS-binding mAbs used a diverse repertoire but targeted the RBS epitope similarly and made extensive contacts with the major antigenic site Sb. Together, our data indicate that RBS- and lateral patch-targeting clones are abundant within the human memory B cell pool, and universal vaccine strategies should aim to drive antibodies against both conserved head and stalk epitopes.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
References
-
- Ekiert DC, Kashyap AK, Steel J, Rubrum A, Bhabha G, Khayat R, Lee JH, Dillon MA, O’Neil RE, Faynboym AM, Horowitz M, Horowitz L, Ward AB, Palese P, Webby R, Lerner RA, Bhatt RR, Wilson IA, Cross-neutralization of influenza A viruses mediated by a single antibody loop. Nature 489, 526–532 (2012); published online EpubSep 27 ( 10.1038/nature11414). - DOI - PMC - PubMed
-
- Whittle JR, Zhang R, Khurana S, King LR, Manischewitz J, Golding H, Dormitzer PR, Haynes BF, Walter EB, Moody MA, Kepler TB, Liao HX, Harrison SC, Broadly neutralizing human antibody that recognizes the receptor-binding pocket of influenza virus hemagglutinin. Proc Natl Acad Sci U S A 108, 14216–14221 (2011); published online EpubAug 23 ( 10.1073/pnas.1111497108). - DOI - PMC - PubMed
-
- Raymond DD, Bajic G, Ferdman J, Suphaphiphat P, Settembre EC, Moody MA, Schmidt AG, Harrison SC, Conserved epitope on influenza-virus hemagglutinin head defined by a vaccine-induced antibody. Proc Natl Acad Sci U S A 115, 168–173 (2018); published online EpubJan 2 ( 10.1073/pnas.1715471115). - DOI - PMC - PubMed
-
- Linderman SL, Chambers BS, Zost SJ, Parkhouse K, Li Y, Herrmann C, Ellebedy AH, Carter DM, Andrews SF, Zheng NY, Huang M, Huang Y, Strauss D, Shaz BH, Hodinka RL, Reyes-Teran G, Ross TM, Wilson PC, Ahmed R, Bloom JD, Hensley SE, Potential antigenic explanation for atypical H1N1 infections among middle-aged adults during the 2013–2014 influenza season. Proc Natl Acad Sci U S A 111, 15798–15803 (2014); published online EpubNov 4 ( 10.1073/pnas.1409171111). - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- U19 AI057266/AI/NIAID NIH HHS/United States
- 75N93019C00028/AI/NIAID NIH HHS/United States
- R01 GM129325/GM/NIGMS NIH HHS/United States
- U19 AI109946/AI/NIAID NIH HHS/United States
- HHSN272201400005C/AI/NIAID NIH HHS/United States
- 75N93019C00051/AI/NIAID NIH HHS/United States
- U19 AI082724/AI/NIAID NIH HHS/United States
- T32 HL007605/HL/NHLBI NIH HHS/United States
- P41 GM103311/GM/NIGMS NIH HHS/United States
- HHSN272201400008C/AI/NIAID NIH HHS/United States
- T32 AI007244/AI/NIAID NIH HHS/United States
- R01 AI145870/AI/NIAID NIH HHS/United States
- K99 AI159136/AI/NIAID NIH HHS/United States
- P01 AI097092/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
