

Engineered reproductively isolated species drive reversible population replacement
- PMID: 34078888
- PMCID: PMC8173020
- DOI: 10.1038/s41467-021-23531-z
Engineered reproductively isolated species drive reversible population replacement
Abstract
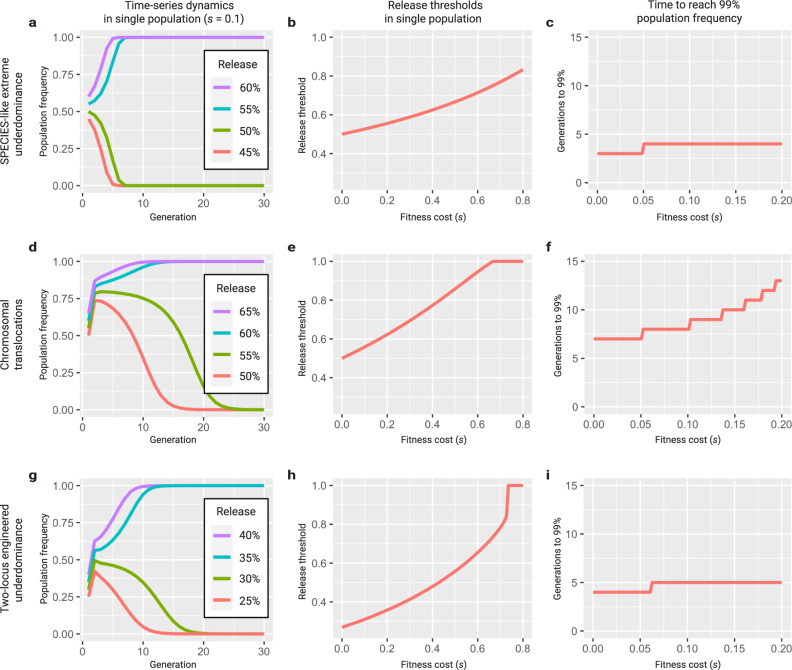
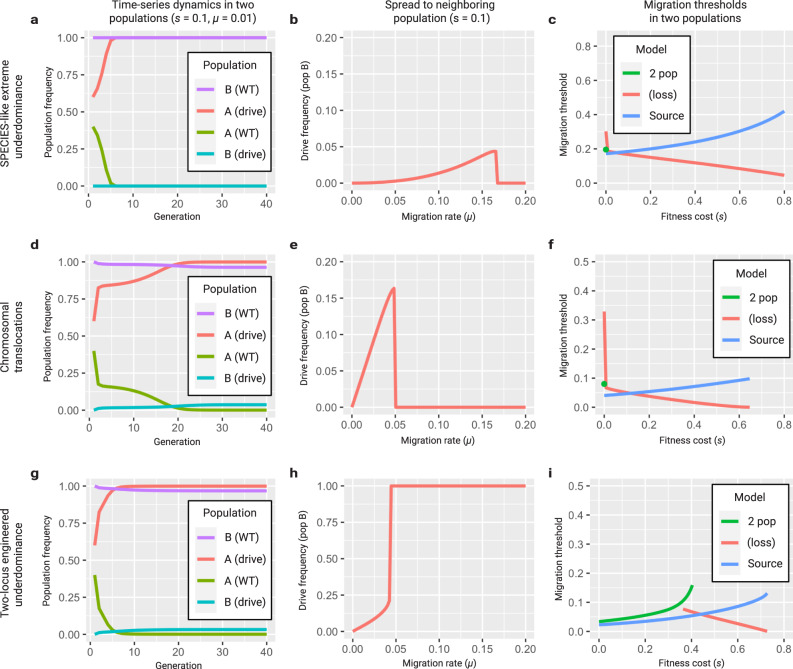
Engineered reproductive species barriers are useful for impeding gene flow and driving desirable genes into wild populations in a reversible threshold-dependent manner. However, methods to generate synthetic barriers are lacking in advanced eukaryotes. Here, to overcome this challenge, we engineer SPECIES (Synthetic Postzygotic barriers Exploiting CRISPR-based Incompatibilities for Engineering Species), an engineered genetic incompatibility approach, to generate postzygotic reproductive barriers. Using this approach, we create multiple reproductively isolated SPECIES and demonstrate their reproductive isolation and threshold-dependent gene drive capabilities in D. melanogaster. Given the near-universal functionality of CRISPR tools, this approach should be portable to many species, including insect disease vectors in which confinable gene drives could be of great practical utility.
Conflict of interest statement
O.S.A. and A.B. have a patent pending on this technology. Patent applicant—University of California, UCSD. Name of inventor(s)—A.B. and O.S.A. Application number—Invention 2020-184. Status of application—Pending. Specific aspect of manuscript covered in patent application—Using multiplexed dCas9/Cas9/gRNA combinations to generate reproductive isolation that can be used for population control. O.S.A. is co-founder and serves on the scientific advisory board of Agragene. All other authors declare no significant competing financial, professional, or personal interests that might have influenced the performance or presentation of the work described.
Figures
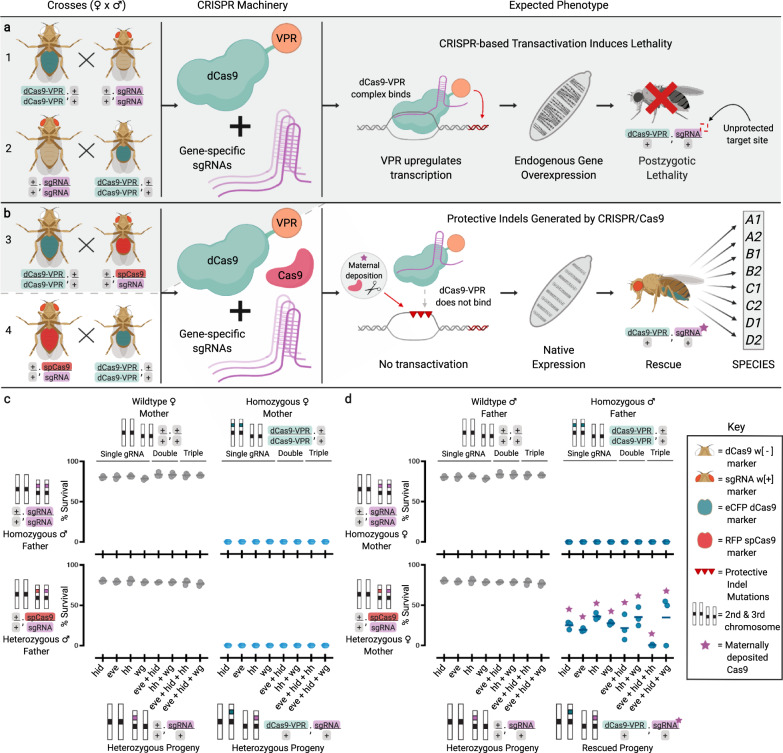
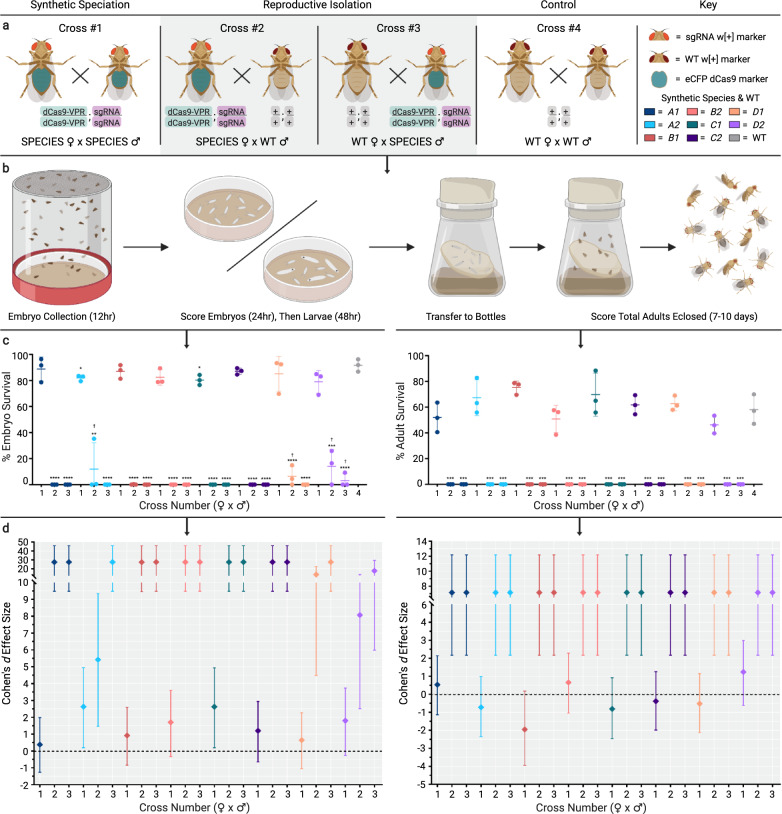
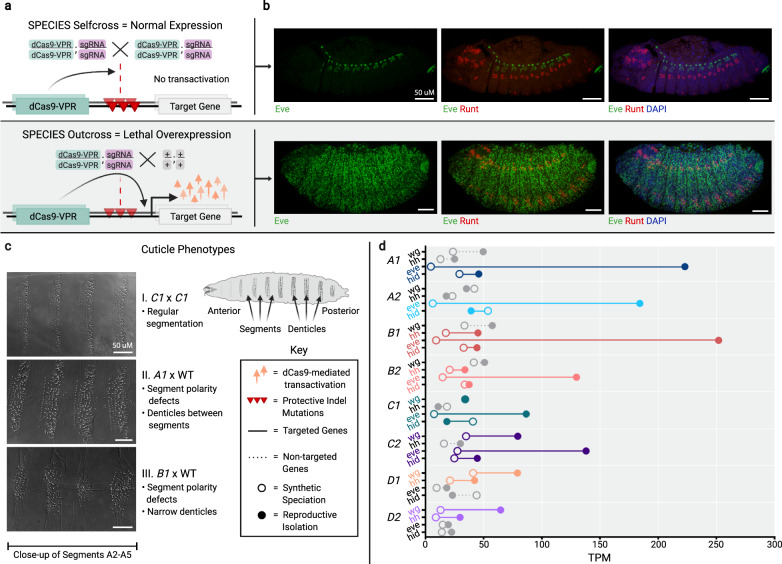
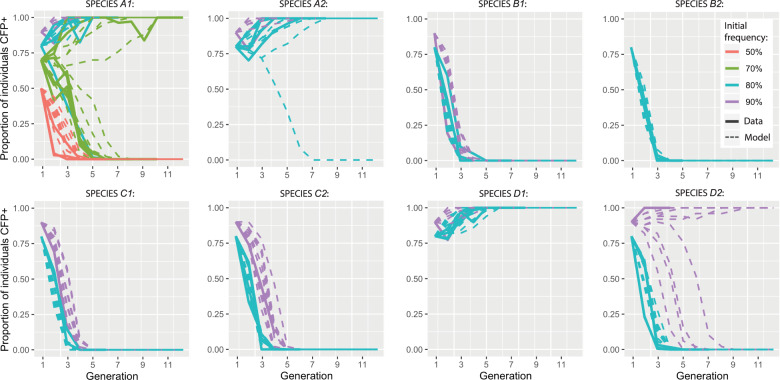
Similar articles
-
Rationally-engineered reproductive barriers using CRISPR & CRISPRa: an evaluation of the synthetic species concept in Drosophila melanogaster.Sci Rep. 2018 Sep 3;8(1):13125. doi: 10.1038/s41598-018-31433-2. Sci Rep. 2018. PMID: 30177778 Free PMC article.
-
Computational and experimental performance of CRISPR homing gene drive strategies with multiplexed gRNAs.Sci Adv. 2020 Mar 4;6(10):eaaz0525. doi: 10.1126/sciadv.aaz0525. eCollection 2020 Mar. Sci Adv. 2020. PMID: 32181354 Free PMC article.
-
Molecular safeguarding of CRISPR gene drive experiments.Elife. 2019 Jan 22;8:e41439. doi: 10.7554/eLife.41439. Elife. 2019. PMID: 30666960 Free PMC article.
-
CRISPR-Based Gene Drives for Pest Control.Trends Biotechnol. 2018 Feb;36(2):130-133. doi: 10.1016/j.tibtech.2017.10.001. Trends Biotechnol. 2018. PMID: 29221716 Review.
-
Concerning RNA-guided gene drives for the alteration of wild populations.Elife. 2014 Jul 17;3:e03401. doi: 10.7554/eLife.03401. Elife. 2014. PMID: 25035423 Free PMC article. Review.
Cited by
-
Genetically engineered insects with sex-selection and genetic incompatibility enable population suppression.Elife. 2022 Feb 2;11:e71230. doi: 10.7554/eLife.71230. Elife. 2022. PMID: 35108195 Free PMC article.
-
Modeling-informed Engineered Genetic Incompatibility strategies to overcome resistance in the invasive Drosophila suzukii.Front Insect Sci. 2022 Nov 22;2:1063789. doi: 10.3389/finsc.2022.1063789. eCollection 2022. Front Insect Sci. 2022. PMID: 38468757 Free PMC article.
-
Predicting thresholds for population replacement gene drives.BMC Biol. 2024 Feb 19;22(1):40. doi: 10.1186/s12915-024-01823-2. BMC Biol. 2024. PMID: 38369493 Free PMC article.
-
Adequacy and sufficiency evaluation of existing EFSA guidelines for the molecular characterisation, environmental risk assessment and post-market environmental monitoring of genetically modified insects containing engineered gene drives.EFSA J. 2020 Nov 12;18(11):e06297. doi: 10.2903/j.efsa.2020.6297. eCollection 2020 Nov. EFSA J. 2020. PMID: 33209154 Free PMC article.
-
Engineering multiple species-like genetic incompatibilities in insects.Nat Commun. 2020 Sep 8;11(1):4468. doi: 10.1038/s41467-020-18348-1. Nat Commun. 2020. PMID: 32901021 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
