

Prohibitin 1 is essential to preserve mitochondria and myelin integrity in Schwann cells
- PMID: 34078899
- PMCID: PMC8172551
- DOI: 10.1038/s41467-021-23552-8
Prohibitin 1 is essential to preserve mitochondria and myelin integrity in Schwann cells
Abstract
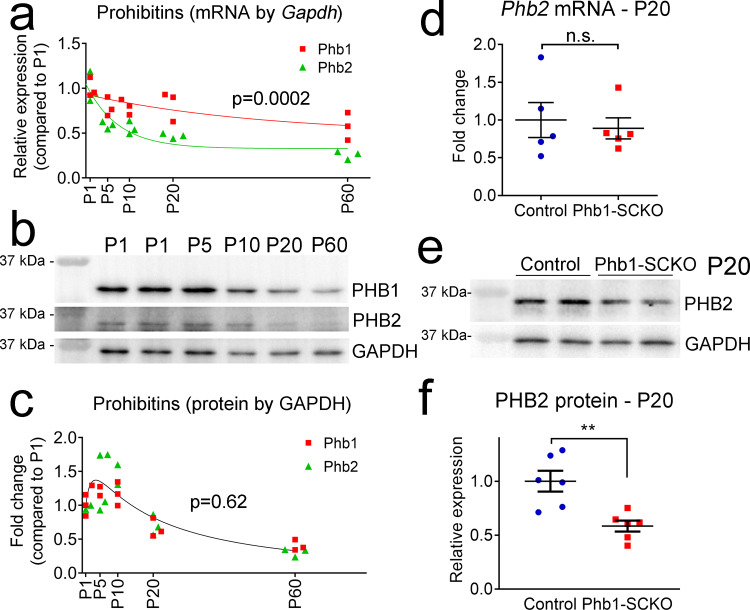
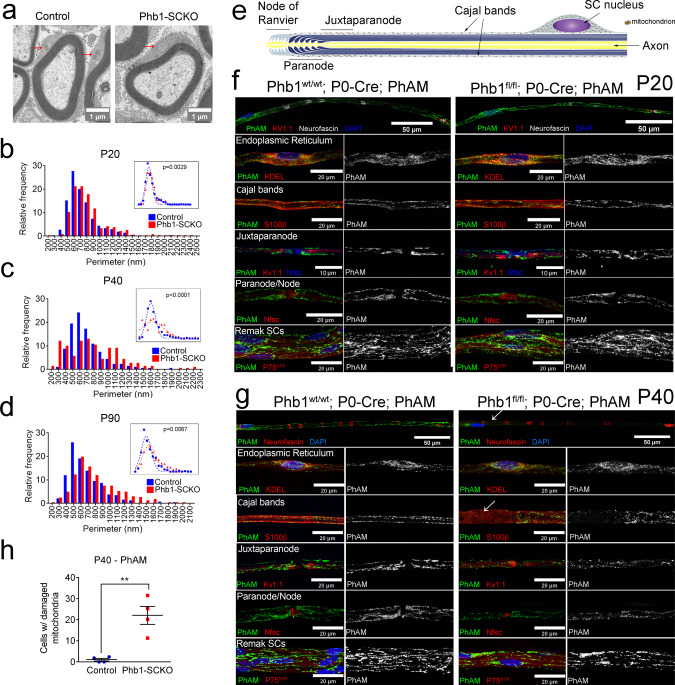
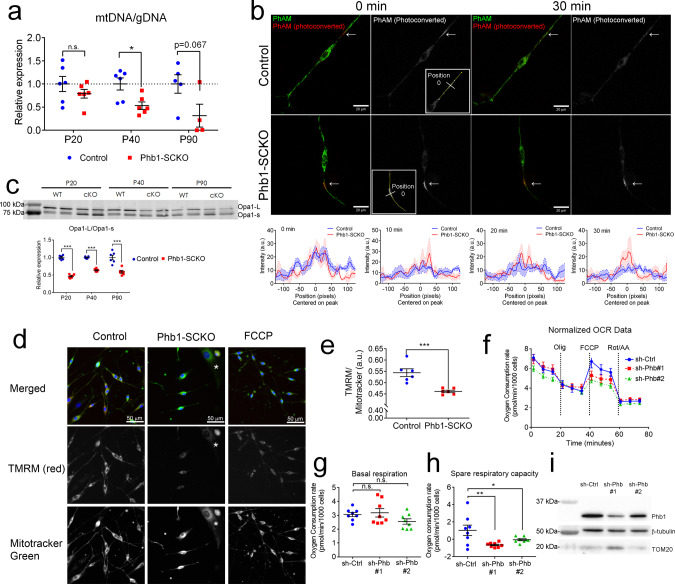
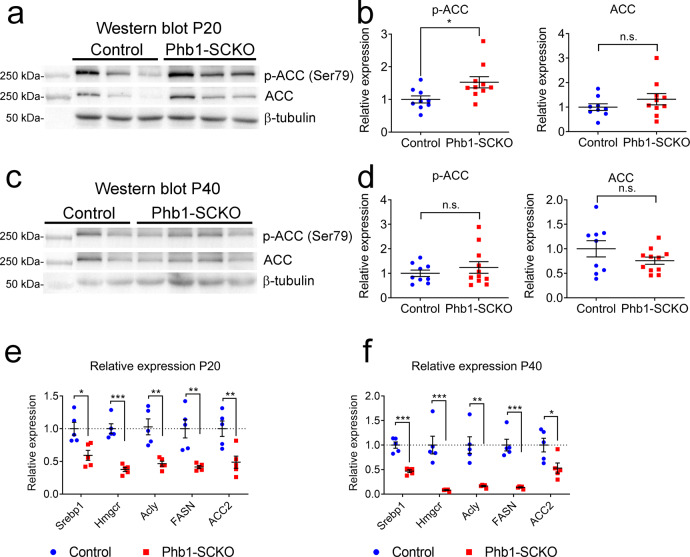
In peripheral nerves, Schwann cells form myelin and provide trophic support to axons. We previously showed that the mitochondrial protein prohibitin 2 can localize to the axon-Schwann-cell interface and is required for developmental myelination. Whether the homologous protein prohibitin 1 has a similar role, and whether prohibitins also play important roles in Schwann cell mitochondria is unknown. Here, we show that deletion of prohibitin 1 in Schwann cells minimally perturbs development, but later triggers a severe demyelinating peripheral neuropathy. Moreover, mitochondria are heavily affected by ablation of prohibitin 1 and demyelination occurs preferentially in cells with apparent mitochondrial loss. Furthermore, in response to mitochondrial damage, Schwann cells trigger the integrated stress response, but, contrary to what was previously suggested, this response is not detrimental in this context. These results identify a role for prohibitin 1 in myelin integrity and advance our understanding about the Schwann cell response to mitochondrial damage.
Conflict of interest statement
The authors declare no competing interests.
Figures



Similar articles
-
Loss of prohibitin 2 in Schwann cells dysregulates key transcription factors controlling developmental myelination.Glia. 2024 Dec;72(12):2247-2267. doi: 10.1002/glia.24610. Epub 2024 Aug 31. Glia. 2024. PMID: 39215540
-
Activation of mTORC1 and c-Jun by Prohibitin1 loss in Schwann cells may link mitochondrial dysfunction to demyelination.Elife. 2021 Sep 14;10:e66278. doi: 10.7554/eLife.66278. Elife. 2021. PMID: 34519641 Free PMC article.
-
Mitochondrial schwannopathy and peripheral myelinopathy in a rabbit model of dideoxycytidine neurotoxicity.Lab Invest. 1994 May;70(5):724-39. Lab Invest. 1994. PMID: 7515130
-
Schwann cell mitochondria as key regulators in the development and maintenance of peripheral nerve axons.Cell Mol Life Sci. 2017 Mar;74(5):827-835. doi: 10.1007/s00018-016-2364-1. Epub 2016 Sep 16. Cell Mol Life Sci. 2017. PMID: 27638763 Free PMC article. Review.
-
Role of the Schwann cell in diabetic neuropathy.Int Rev Neurobiol. 2002;50:293-321. doi: 10.1016/s0074-7742(02)50081-7. Int Rev Neurobiol. 2002. PMID: 12198814 Review.
Cited by
-
Subcellular distribution of prohibitin 1 in rat liver during liver regeneration and its cellular implication.World J Hepatol. 2024 Jan 27;16(1):65-74. doi: 10.4254/wjh.v16.i1.65. World J Hepatol. 2024. PMID: 38313239 Free PMC article.
-
Loss of prohibitin 2 in Schwann cells dysregulates key transcription factors controlling developmental myelination.Glia. 2024 Dec;72(12):2247-2267. doi: 10.1002/glia.24610. Epub 2024 Aug 31. Glia. 2024. PMID: 39215540
-
Peripheral glia diversity.J Anat. 2022 Nov;241(5):1219-1234. doi: 10.1111/joa.13484. Epub 2021 Jun 15. J Anat. 2022. PMID: 34131911 Free PMC article. Review.
-
Traumatic and Diabetic Schwann Cell Demyelination Is Triggered by a Transient Mitochondrial Calcium Release through Voltage Dependent Anion Channel 1.Biomedicines. 2022 Jun 19;10(6):1447. doi: 10.3390/biomedicines10061447. Biomedicines. 2022. PMID: 35740468 Free PMC article.
-
Loss of prohibitin 2 in Schwann cells dysregulates key transcription factors controlling developmental myelination.bioRxiv [Preprint]. 2024 Mar 20:2024.03.20.585915. doi: 10.1101/2024.03.20.585915. bioRxiv. 2024. Update in: Glia. 2024 Dec;72(12):2247-2267. doi: 10.1002/glia.24610. PMID: 38562812 Free PMC article. Updated. Preprint.
