

Metabolic remodeling precedes mTORC1-mediated cardiac hypertrophy
- PMID: 34081952
- PMCID: PMC8630806
- DOI: 10.1016/j.yjmcc.2021.05.016
Metabolic remodeling precedes mTORC1-mediated cardiac hypertrophy
Abstract
Rationale: The nutrient sensing mechanistic target of rapamycin complex 1 (mTORC1) and its primary inhibitor, tuberin (TSC2), are cues for the development of cardiac hypertrophy. The phenotype of mTORC1 induced hypertrophy is unknown.
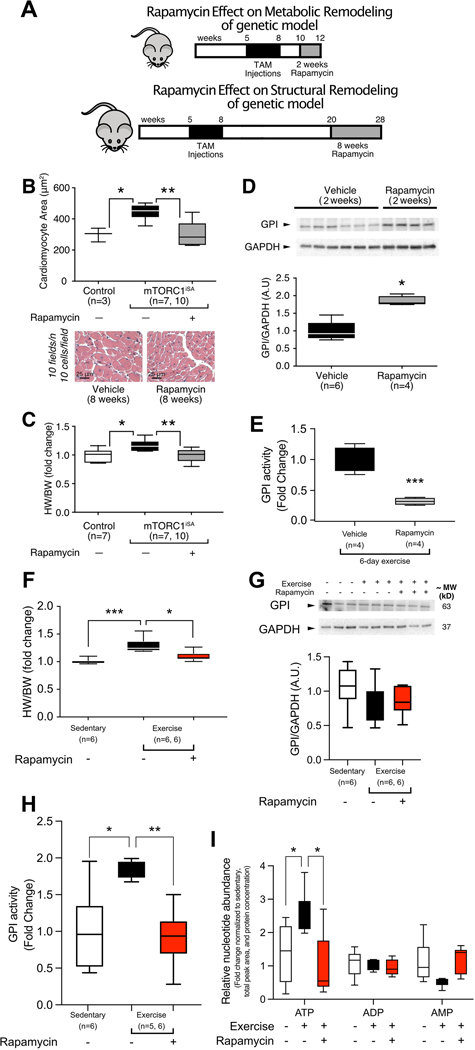
Objective: To examine the impact of sustained mTORC1 activation on metabolism, function, and structure of the adult heart.
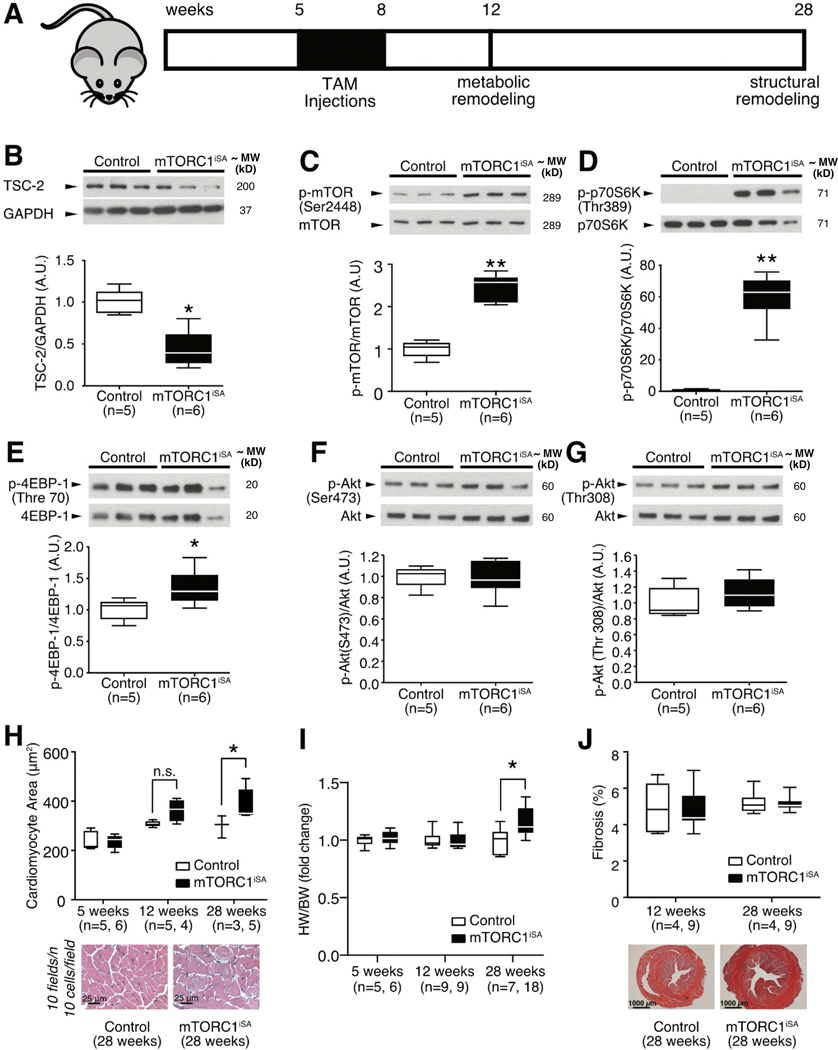
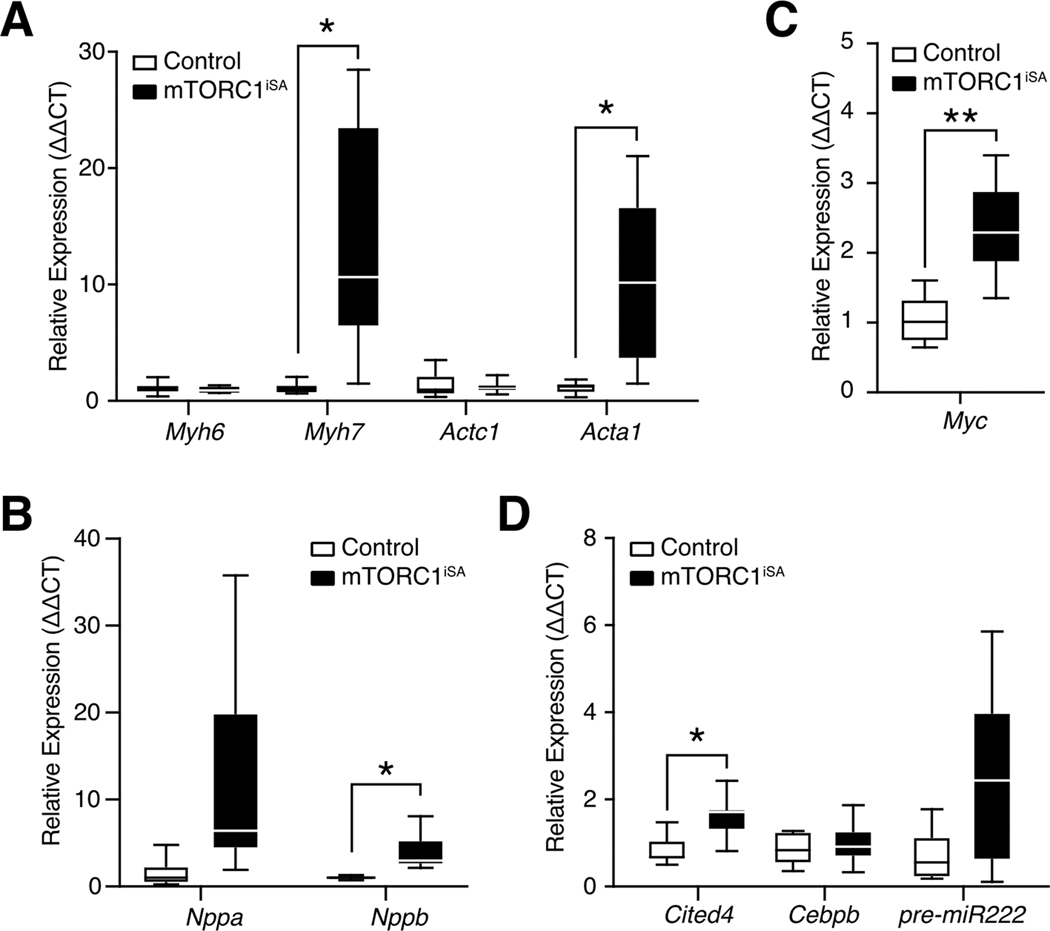
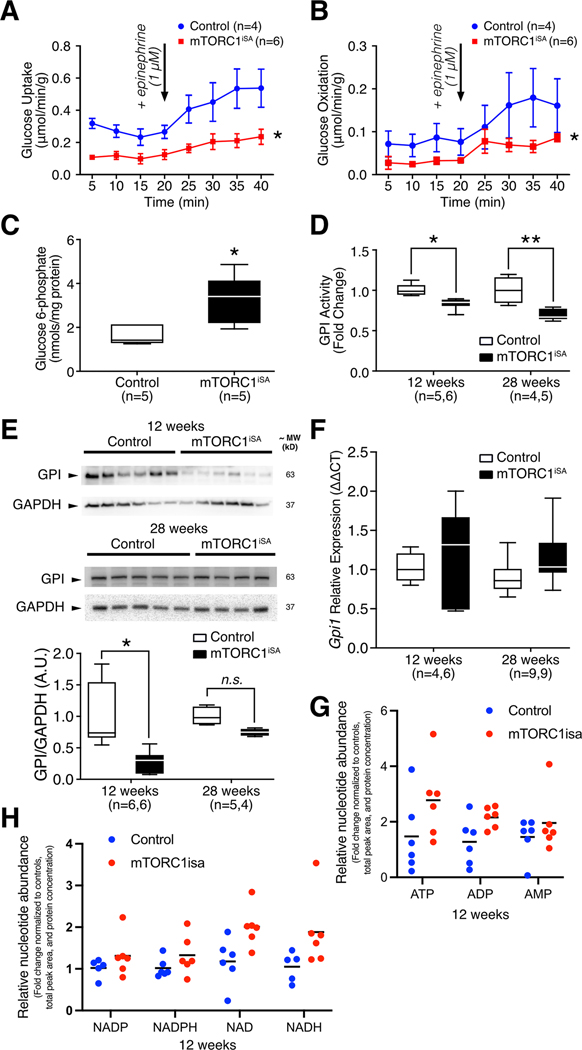
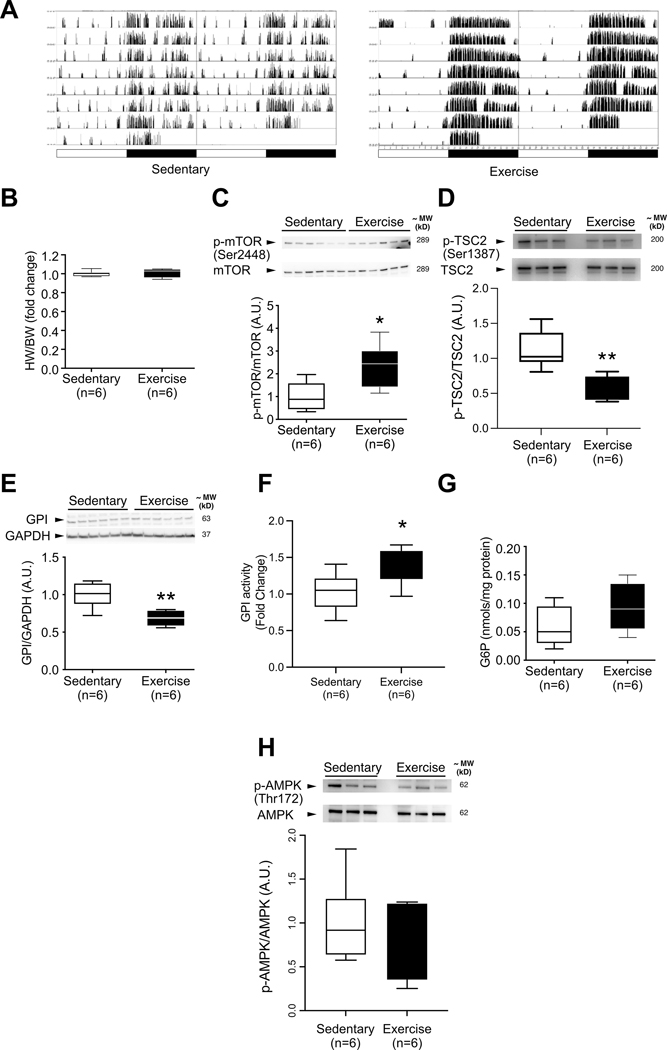
Methods and results: We developed a mouse model of inducible, cardiac-specific sustained mTORC1 activation (mTORC1iSA) through deletion of Tsc2. Prior to hypertrophy, rates of glucose uptake and oxidation, as well as protein and enzymatic activity of glucose 6-phosphate isomerase (GPI) were decreased, while intracellular levels of glucose 6-phosphate (G6P) were increased. Subsequently, hypertrophy developed. Transcript levels of the fetal gene program and pathways of exercise-induced hypertrophy increased, while hypertrophy did not progress to heart failure. We therefore examined the hearts of wild-type mice subjected to voluntary physical activity and observed early changes in GPI, followed by hypertrophy. Rapamycin prevented these changes in both models.
Conclusion: Activation of mTORC1 in the adult heart triggers the development of a non-specific form of hypertrophy which is preceded by changes in cardiac glucose metabolism.
Keywords: Exercise; Glycolysis; Hypertrophy; Metabolism; mTORC1.
Copyright © 2021 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Disclosures
The authors have no conflict of interests to disclose.
Figures
References
-
- Meerson FZ, Kapelko VI, The contractile function of the myocardium in two types of cardiac adaptation to a chronic load, Cardiology 57(4) (1972) 183–99. - PubMed
-
- Hill JA, Olson EN, Cardiac plasticity, N Engl J Med 358(13) (2008) 1370–80. - PubMed
-
- van Bilsen M, van Nieuwenhoven FA, van der Vusse GJ, Metabolic remodelling of the failing heart: beneficial or detrimental?, Cardiovasc Res 81(3) (2009) 420–8. - PubMed
-
- Saxton RA, Sabatini DM, mTOR Signaling in Growth, Metabolism, and Disease, Cell 169(2) (2017) 361–371. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
