

Hydrostatic mechanical stress regulates growth and maturation of the atrioventricular valve
- PMID: 34086041
- PMCID: PMC8276982
- DOI: 10.1242/dev.196519
Hydrostatic mechanical stress regulates growth and maturation of the atrioventricular valve
Abstract
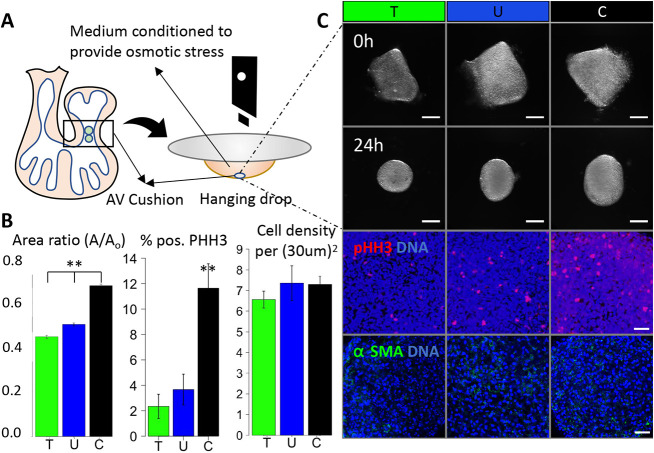
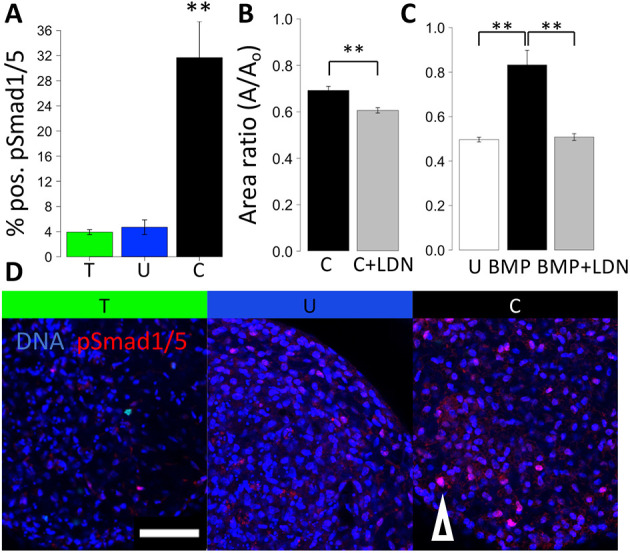
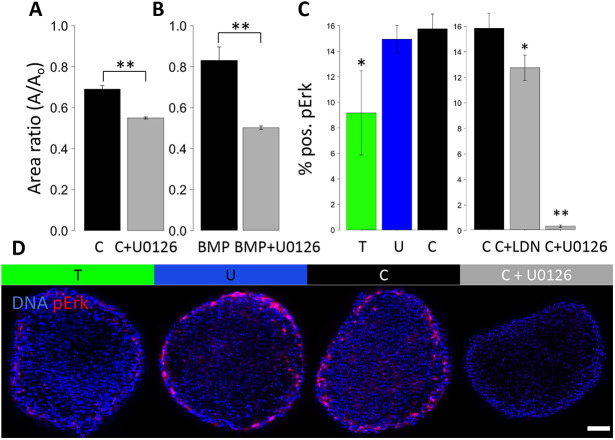
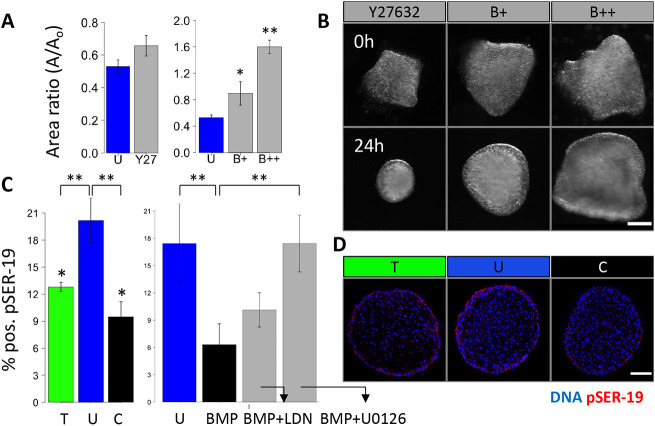
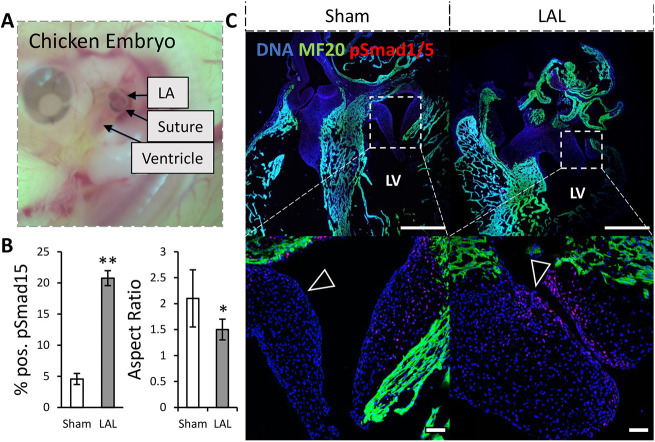
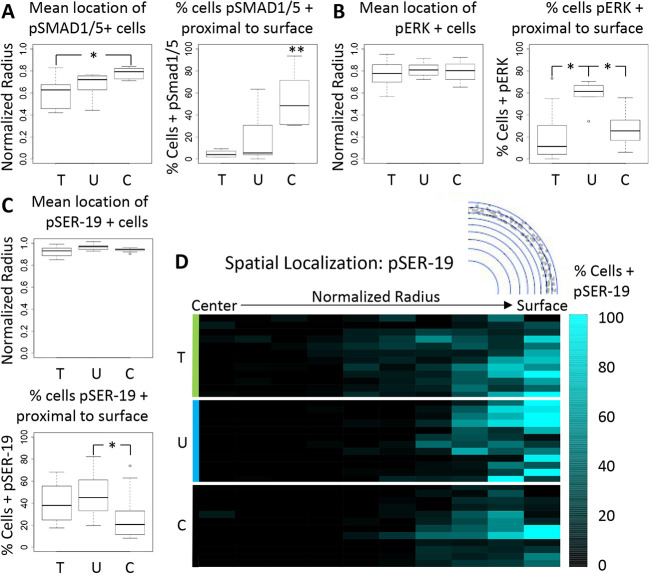
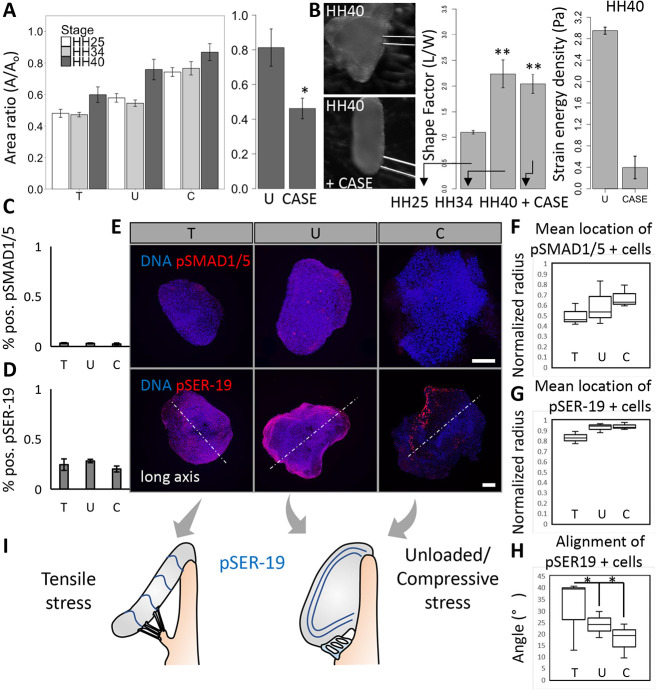
During valvulogenesis, cytoskeletal, secretory and transcriptional events drive endocardial cushion growth and remodeling into thin fibrous leaflets. Genetic disorders play an important role in understanding valve malformations but only account for a minority of clinical cases. Mechanical forces are ever present, but how they coordinate molecular and cellular decisions remains unclear. In this study, we used osmotic pressure to interrogate how compressive and tensile stresses influence valve growth and shape maturation. We found that compressive stress drives a growth phenotype, whereas tensile stress increases compaction. We identified a mechanically activated switch between valve growth and maturation, by which compression induces cushion growth via BMP-pSMAD1/5, while tension induces maturation via pSer-19-mediated MLC2 contractility. The compressive stress acts through BMP signaling to increase cell proliferation and decrease cell contractility, and MEK-ERK is essential for both compressive stress and BMP mediation of compaction. We further showed that the effects of osmotic stress are conserved through the condensation and elongation stages of development. Together, our results demonstrate that compressive/tensile stress regulation of BMP-pSMAD1/5 and MLC2 contractility orchestrates valve growth and remodeling.
Keywords: BMP signaling; Cardiac valve development; MLC contractility; Mechanobiology.
© 2021. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interests The authors declare no competing or financial interests.
Figures
References
-
- Aikawa, E., Whittaker, P., Farber, M., Mendelson, K., Padera, R. F., Aikawa, M. and Schoen, F. J. (2006). Human semilunar cardiac valve remodeling by activated cells from fetus to adult: implications for postnatal adaptation, pathology, and tissue engineering. Circulation 113, 1344-1352. 10.1161/CIRCULATIONAHA.105.591768 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
