

NF-κB Rel subunit exchange on a physiological timescale
- PMID: 34089216
- PMCID: PMC8376415
- DOI: 10.1002/pro.4134
NF-κB Rel subunit exchange on a physiological timescale
Abstract
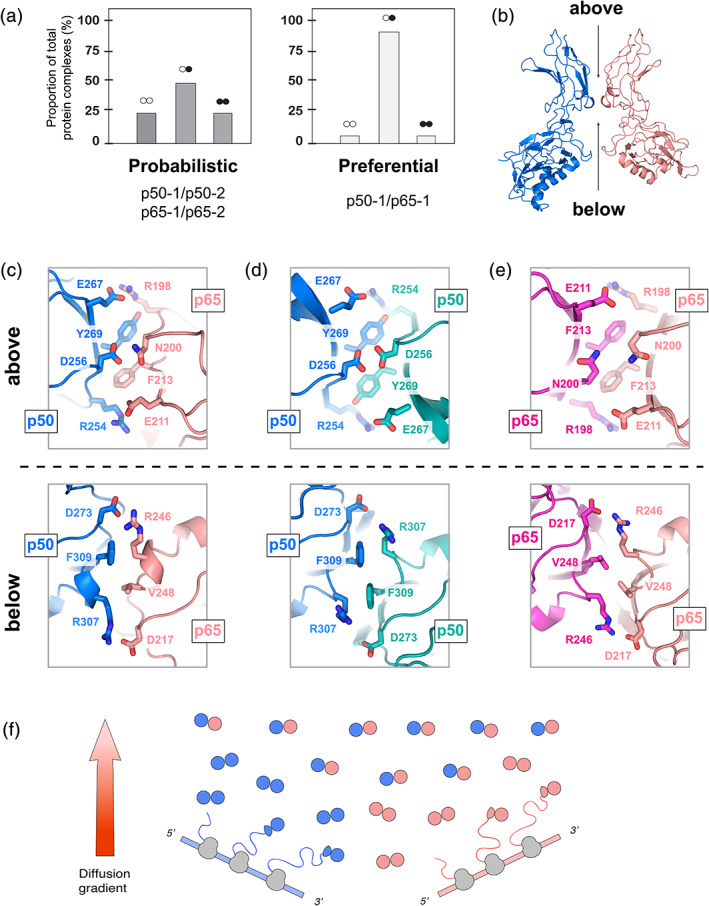
The Rel proteins of the NF-κB complex comprise one of the most investigated transcription factor families, forming a variety of hetero- or homodimers. Nevertheless, very little is known about the fundamental kinetics of NF-κB complex assembly, or the inter-conversion potential of dimerised Rel subunits. Here, we examined an unexplored aspect of NF-κB dynamics, focusing on the dissociation and reassociation of the canonical p50 and p65 Rel subunits and their ability to form new hetero- or homodimers. We employed a soluble expression system to enable the facile production of NF-κB Rel subunits, and verified these proteins display canonical NF-κB nucleic acid binding properties. Using a combination of biophysical techniques, we demonstrated that, at physiological temperatures, homodimeric Rel complexes routinely exchange subunits with a half-life of less than 10 min. In contrast, we found a dramatic preference for the formation of the p50/p65 heterodimer, which demonstrated a kinetic stability of at least an order of magnitude greater than either homodimer. These results suggest that specific DNA targets of either the p50 or p65 homodimers can only be targeted when these subunits are expressed exclusively, or with the intervention of additional post-translational modifications. Together, this work implies a new model of how cells can modulate NF-κB activity by fine-tuning the relative proportions of the p50 and p65 proteins, as well as their time of expression. This work thus provides a new quantitative interpretation of Rel dimer distribution in the cell, particularly for those who are developing mathematical models of NF-κB activity.
Keywords: DNA binding; NF-kappaB; NF-κB; Rel proteins; native mass spectrometry; protein complex; protein dynamics; protein-protein interaction; subunit exchange; transcription factor.
© 2021 The Protein Society.
Conflict of interest statement
The authors declare no competing or conflicting interests.
Figures
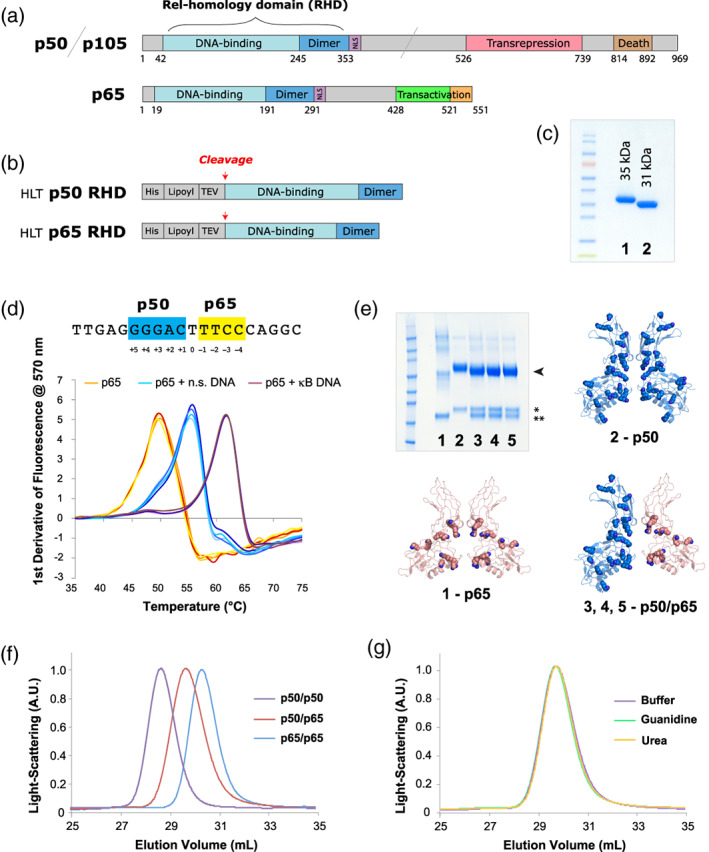
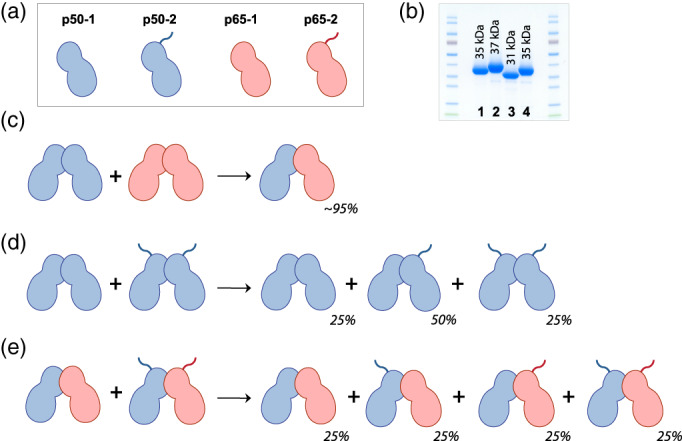
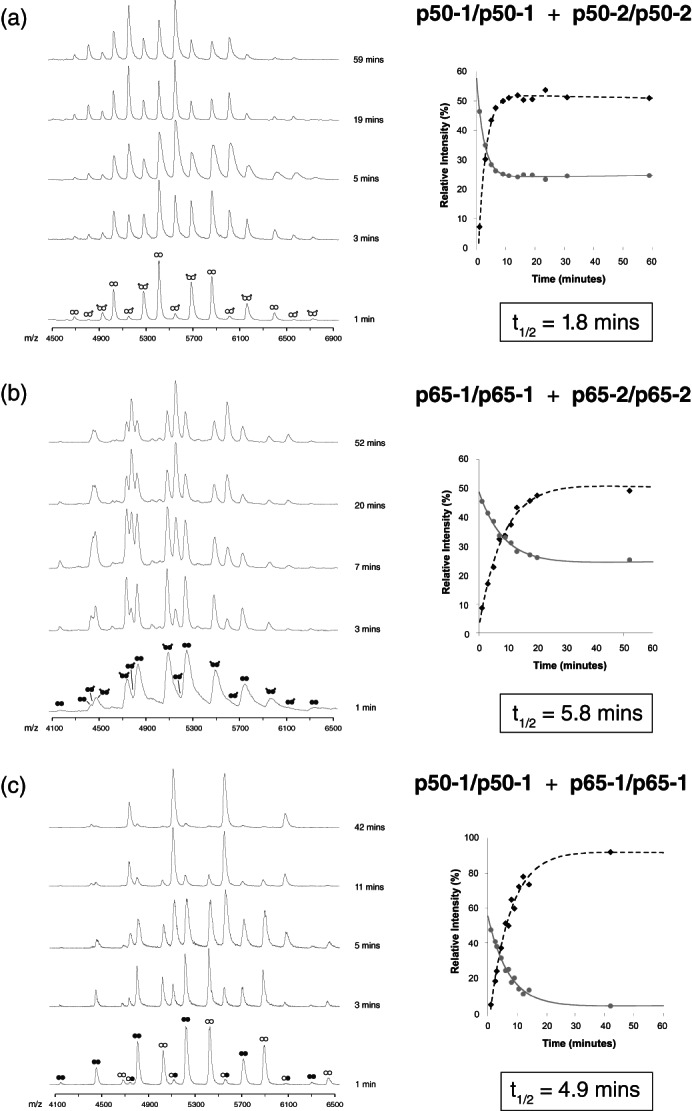
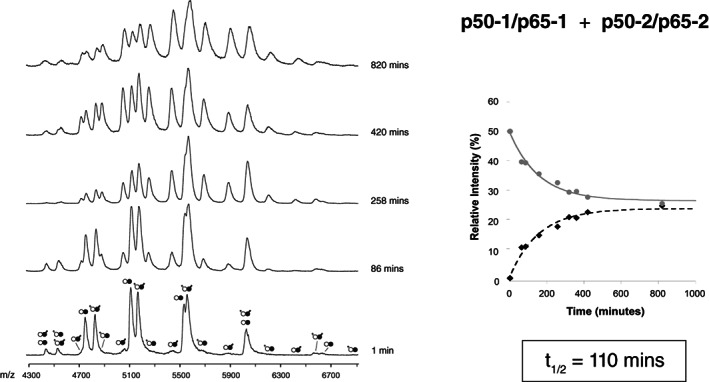
References
-
- Ghosh S. Handbook of transcription factor NF‐kappaB. Boca Raton: CRC Press; 2007.
-
- Gilmore TD. Introduction to NF‐κB: Players, pathways, perspectives. Oncogene. 2006;25:6680–6684. - PubMed
-
- Sen R, Baltimore D. Inducibility of kappa immunoglobulin enhancer‐binding protein Nf‐kappa B by a posttranslational mechanism. Cell. 1986;47:921–928. - PubMed
-
- Sen R, Baltimore D. Multiple nuclear factors interact with the immunoglobulin enhancer sequences. Cell. 1986;46:705–716. - PubMed
-
- Perkins ND. Integrating cell‐signalling pathways with NF‐κB and IKK function. Nat Rev Mol Cell Biol. 2007;8:49–62. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
