

Perturbed hematopoiesis in individuals with germline DNMT3A overgrowth Tatton-Brown-Rahman syndrome
- PMID: 34092059
- PMCID: PMC8968878
- DOI: 10.3324/haematol.2021.278990
Perturbed hematopoiesis in individuals with germline DNMT3A overgrowth Tatton-Brown-Rahman syndrome
Abstract
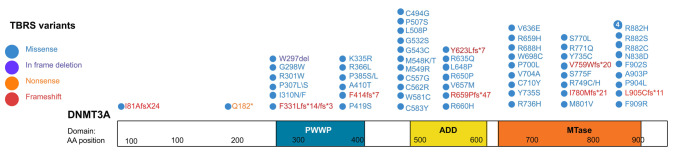
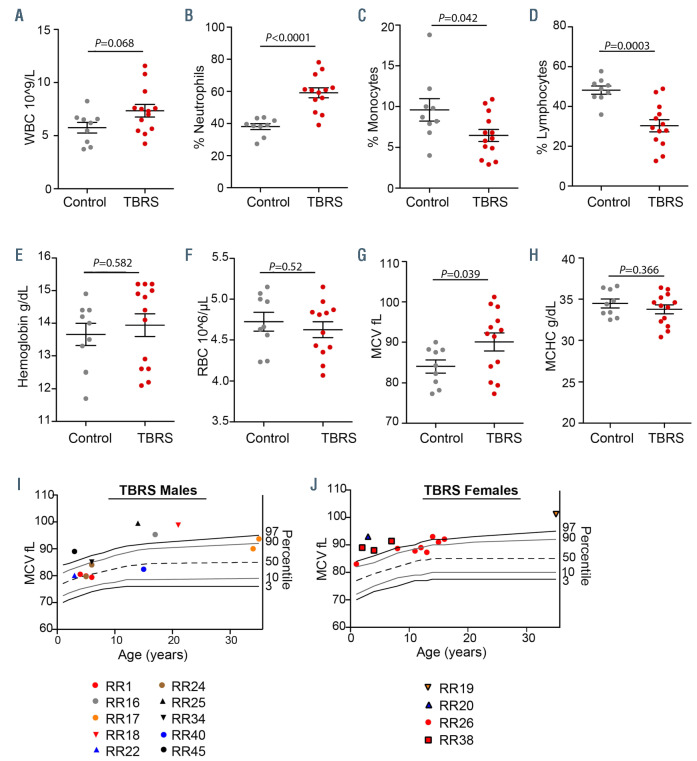
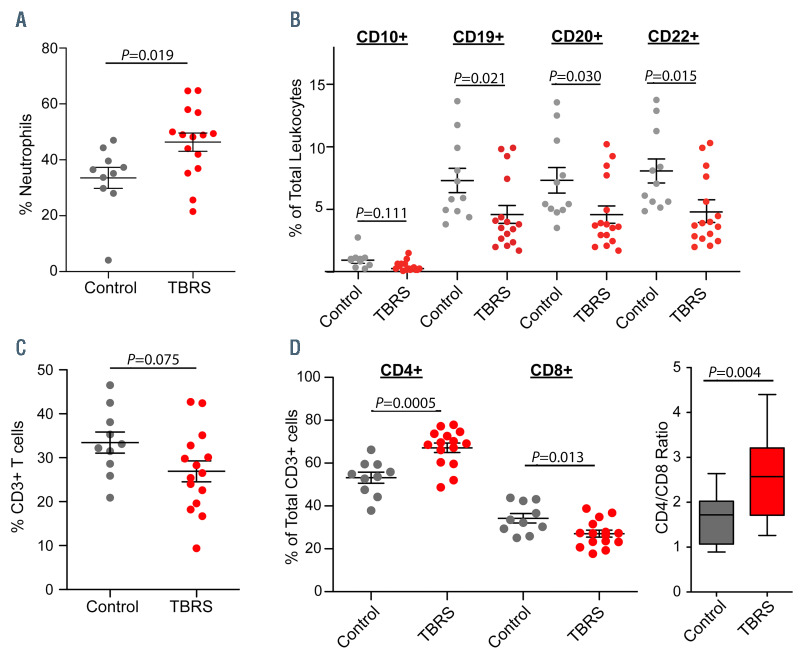
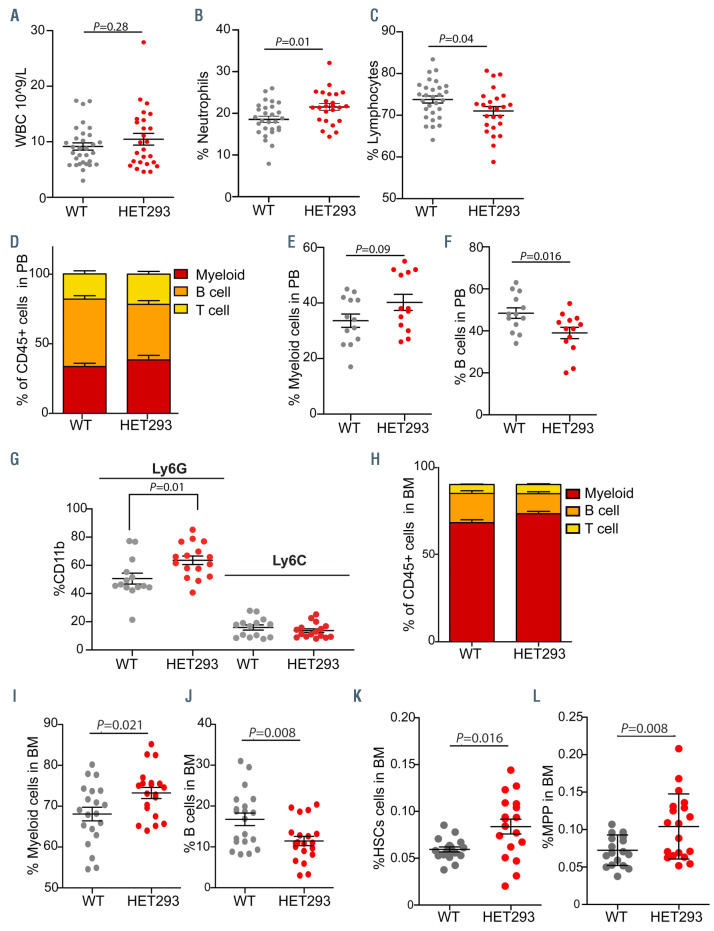
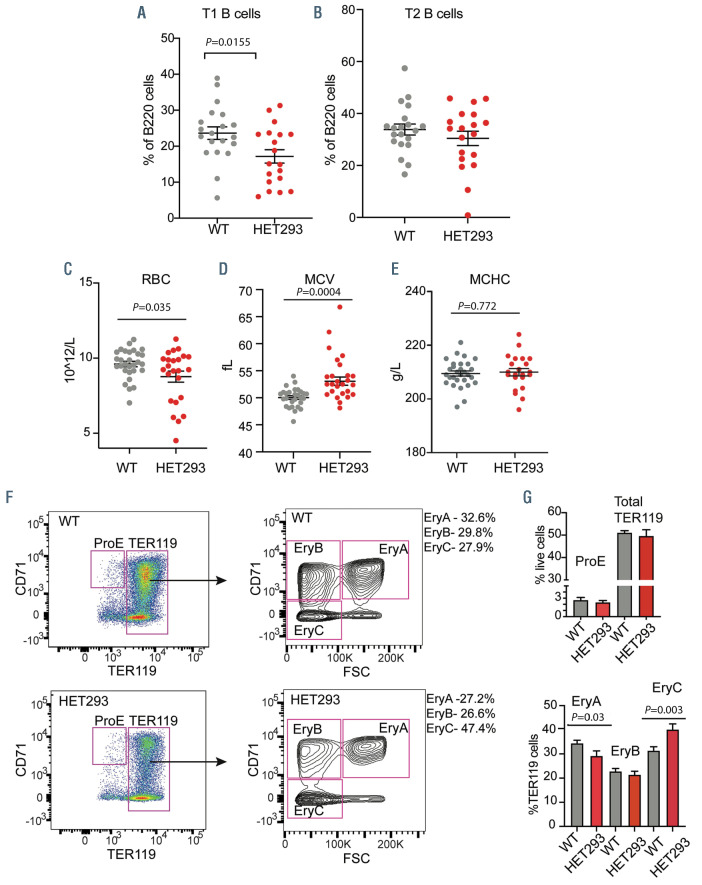
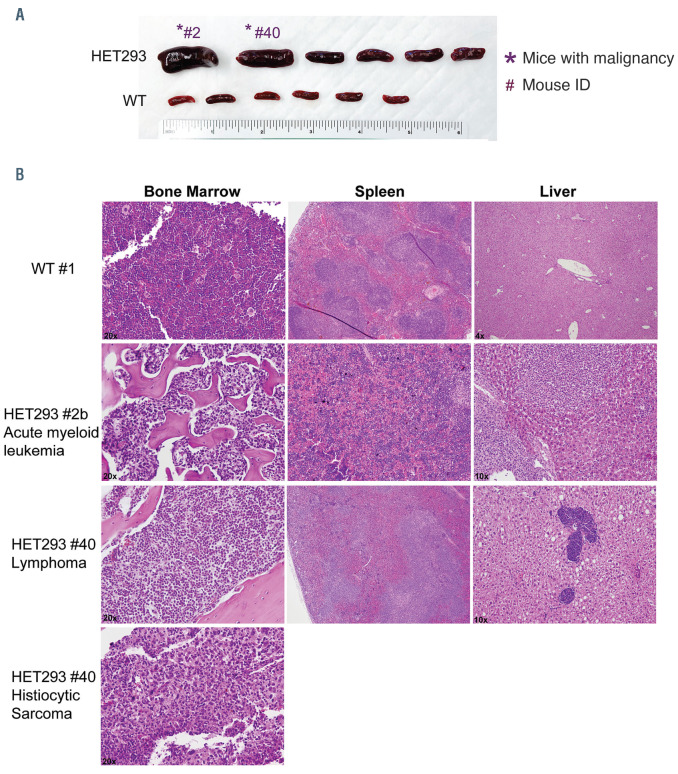
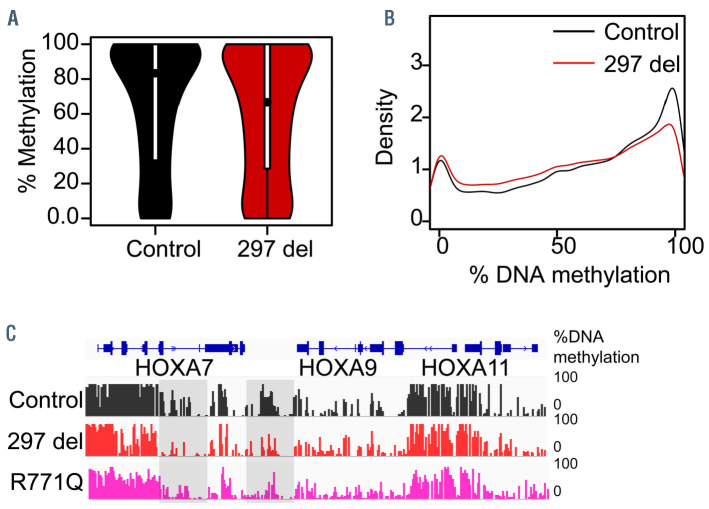
Tatton-Brown-Rahman syndrome (TBRS) is an overgrowth disorder caused by germline heterozygous mutations in the DNA methyltransferase DNMT3A. DNMT3A is a critical regulator of hematopoietic stem cell (HSC) differentiation and somatic DNMT3A mutations are frequent in hematologic malignancies and clonal hematopoiesis. Yet, the impact of constitutive DNMT3A mutation on hematopoiesis in TBRS is undefined. In order to establish how constitutive mutation of DNMT3A impacts blood development in TBRS we gathered clinical data and analyzed blood parameters in 18 individuals with TBRS. We also determined the distribution of major peripheral blood cell lineages by flow cytometric analyses. Our analyses revealed non-anemic macrocytosis, a relative decrease in lymphocytes and increase in neutrophils in TBRS individuals compared to unaffected controls. We were able to recapitulate these hematologic phenotypes in multiple murine models of TBRS and identified rare hematological and non-hematological malignancies associated with constitutive Dnmt3a mutation. We further show that loss of DNMT3A in TBRS is associated with an altered DNA methylation landscape in hematopoietic cells affecting regions critical to stem cell function and tumorigenesis. Overall, our data identify key hematopoietic effects driven by DNMT3A mutation with clinical implications for individuals with TBRS and DNMT3A-associated clonal hematopoiesis or malignancies.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
