

The foraging gene affects alcohol sensitivity, metabolism and memory in Drosophila
- PMID: 34092172
- PMCID: PMC9215342
- DOI: 10.1080/01677063.2021.1931178
The foraging gene affects alcohol sensitivity, metabolism and memory in Drosophila
Abstract
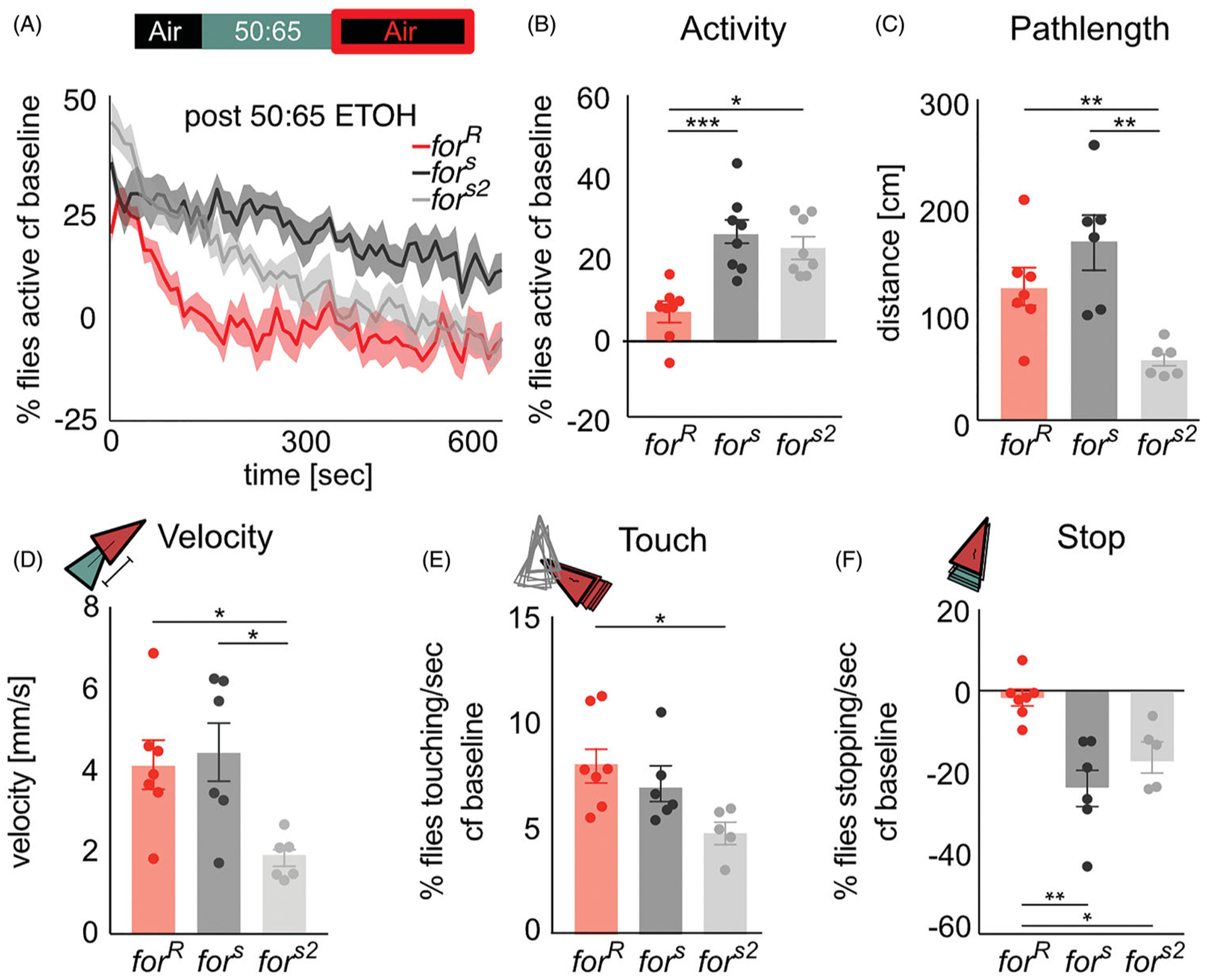
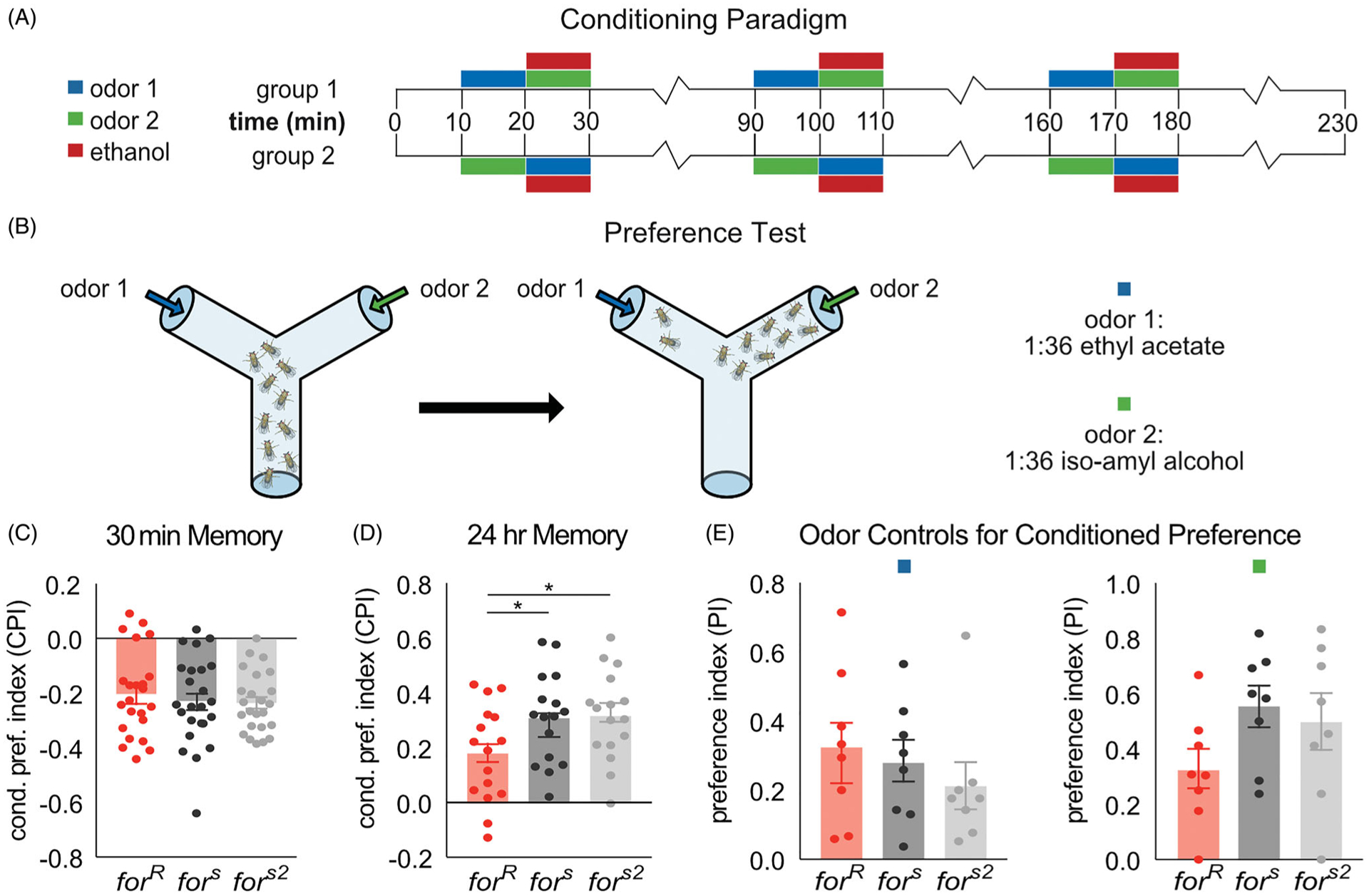
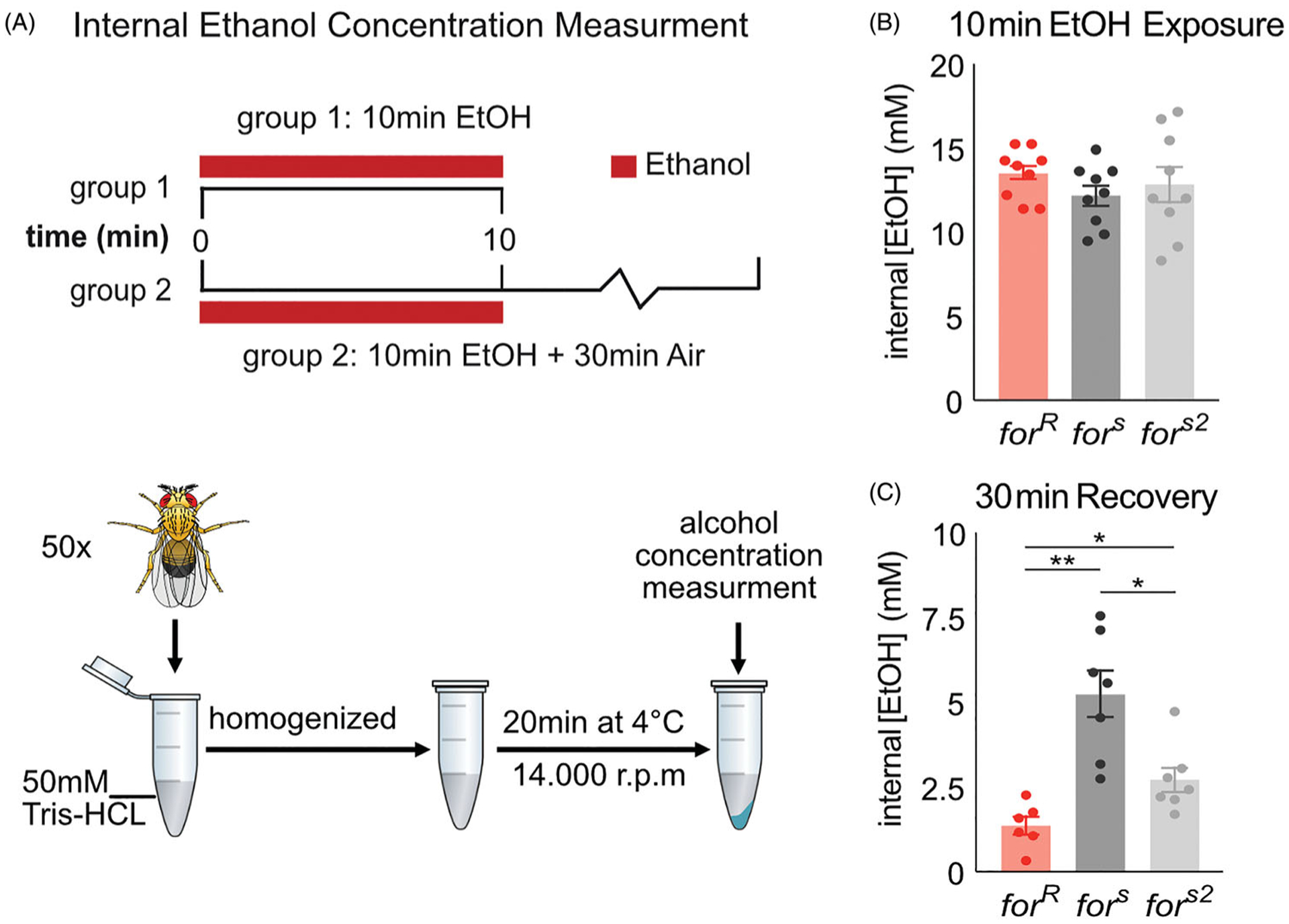
The genetic basis of alcohol use disorder (AUD) is complex. Understanding how natural genetic variation contributes to alcohol phenotypes can help us identify and understand the genetic basis of AUD. Recently, a single nucleotide polymorphism in the human foraging (for) gene ortholog, Protein Kinase cGMP-Dependent 1 (PRKG1), was found to be associated with stress-induced risk for alcohol abuse. However, the mechanistic role that PRKG1 plays in AUD is not well understood. We use natural variation in the Drosophila for gene to describe how variation of cGMP-dependent protein kinase (PKG) activity modifies ethanol-induced phenotypes. We found that variation in for affects ethanol-induced increases in locomotion and memory of the appetitive properties of ethanol intoxication. Further, these differences may stem from the ability to metabolize ethanol. Together, this data suggests that natural variation in PKG modulates cue reactivity for alcohol, and thus could influence alcohol cravings by differentially modulating metabolic and behavioral sensitivities to alcohol.
Keywords: AUD; Drosophila; Foraging; PRKG1; alcohol; cGMP-dependent protein kinase; locomotion; memory; metabolism.
Conflict of interest statement
Disclosure statement
No potential conflict of interest was reported by the author(s).
Figures
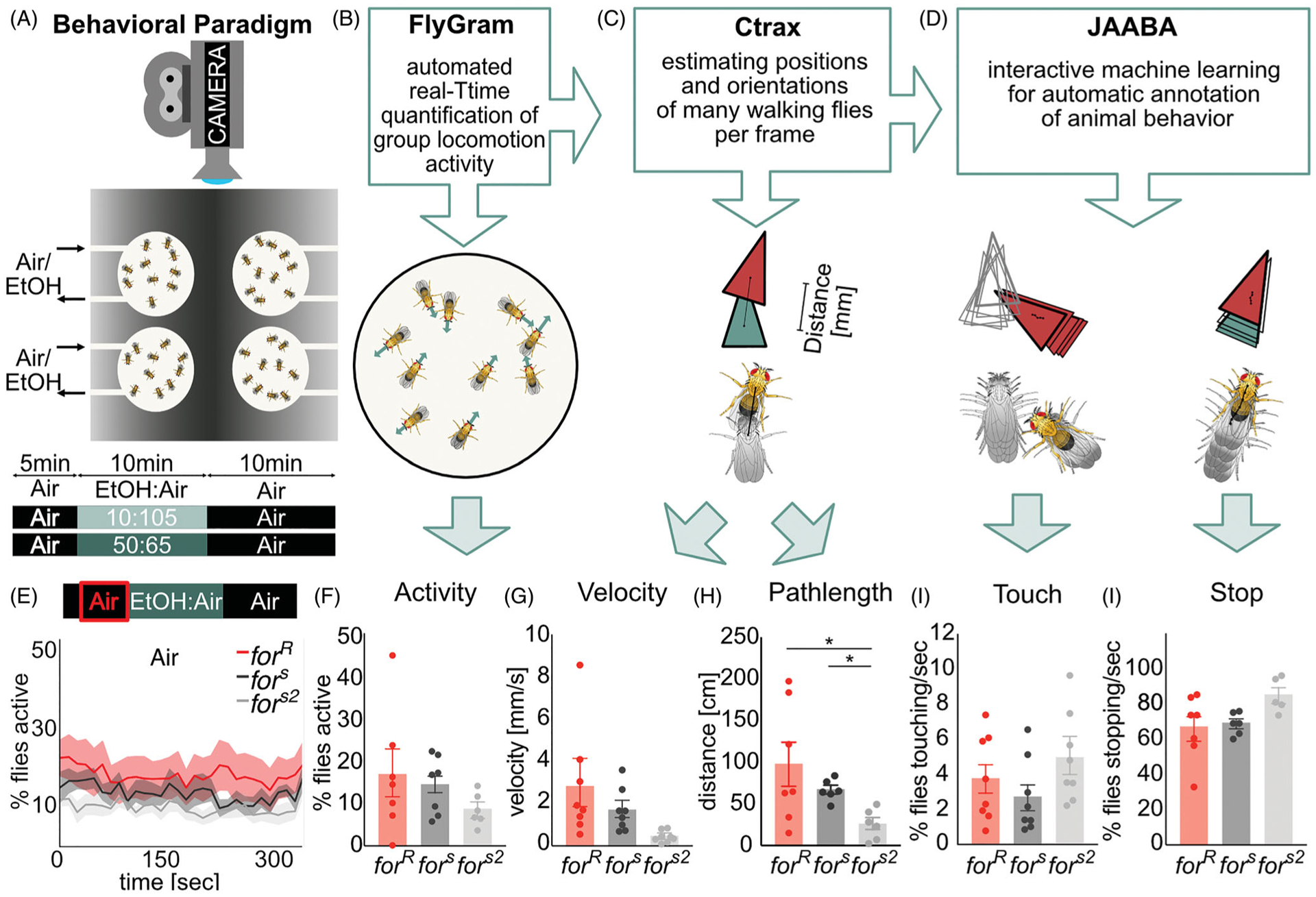
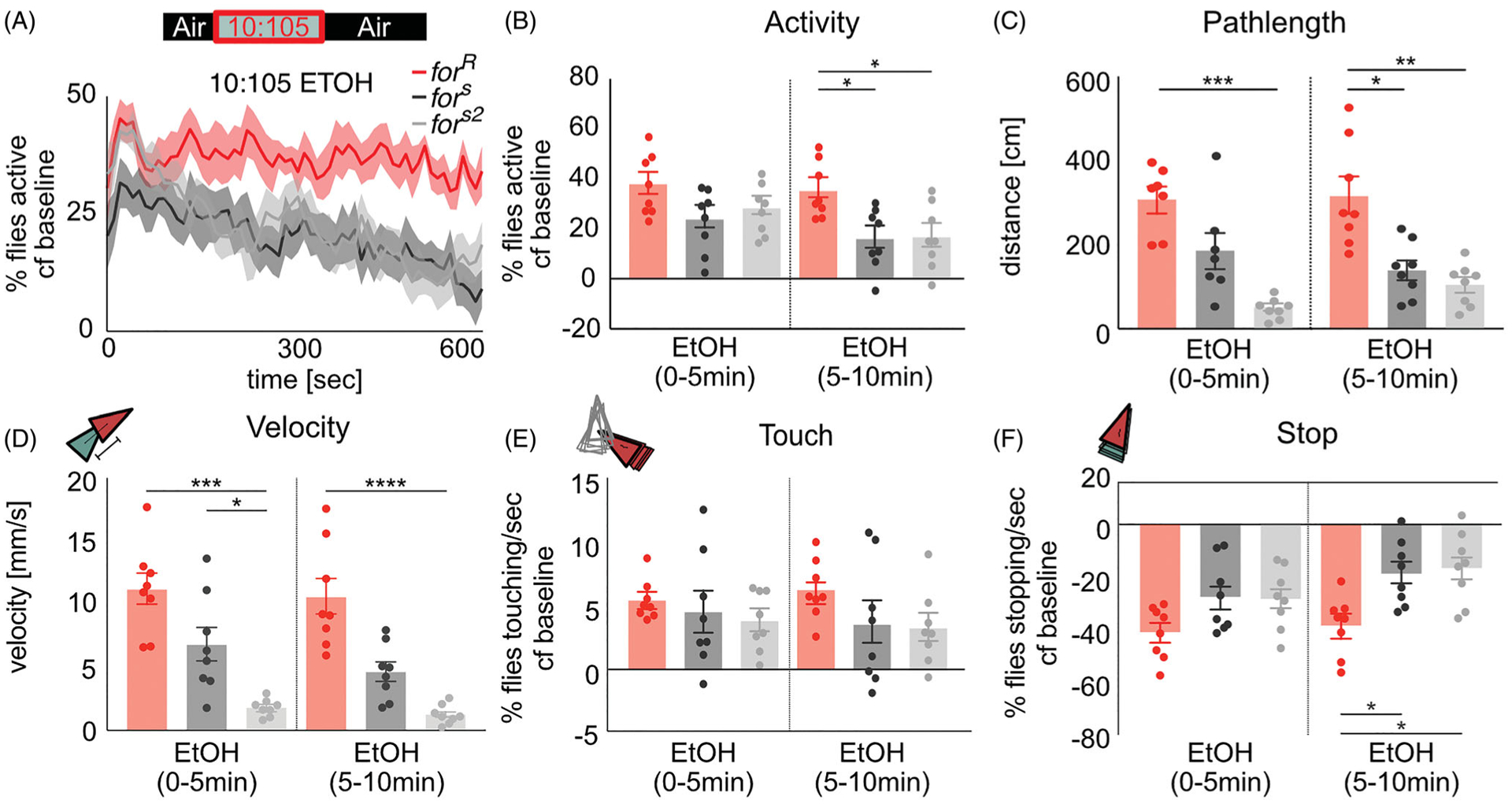
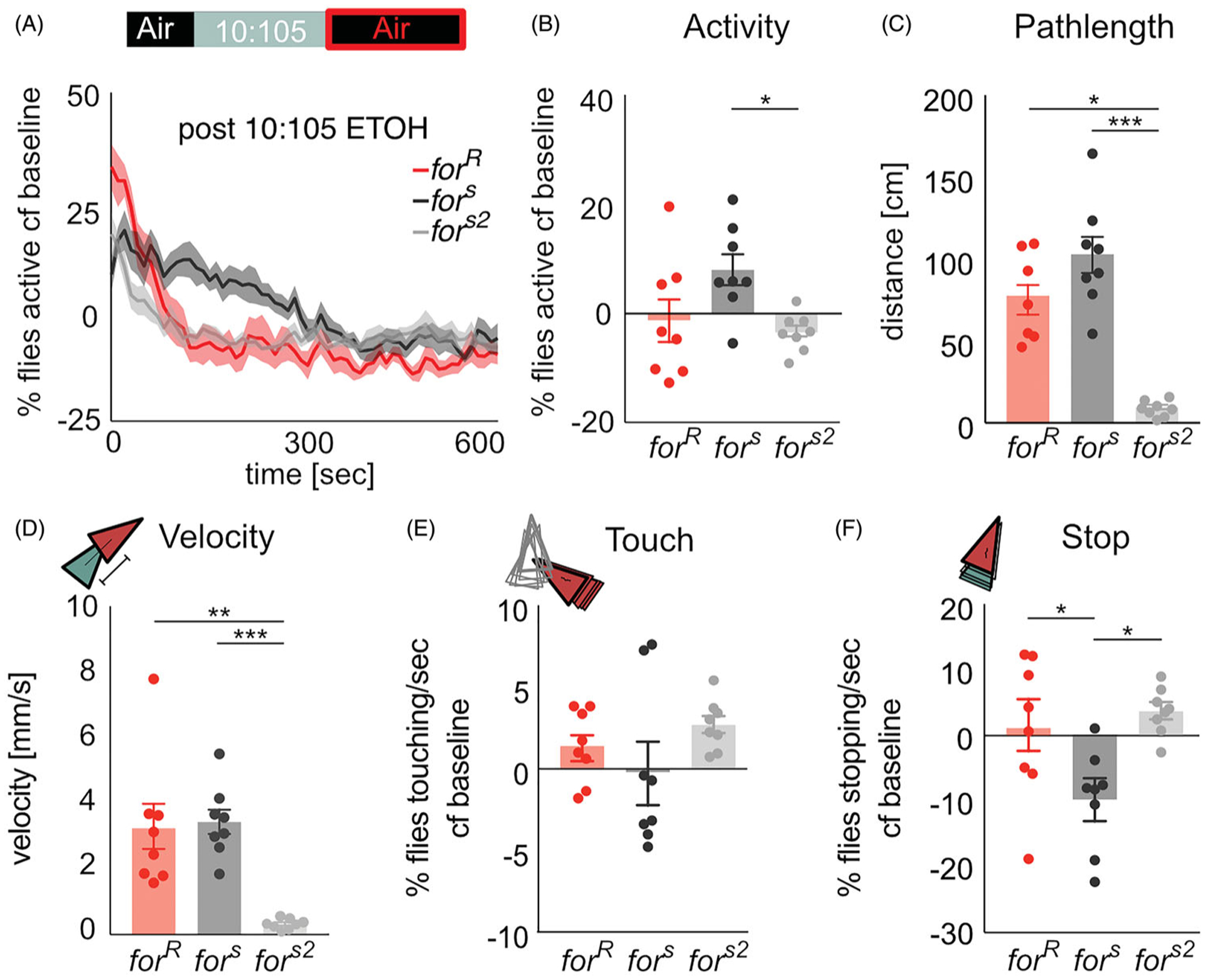
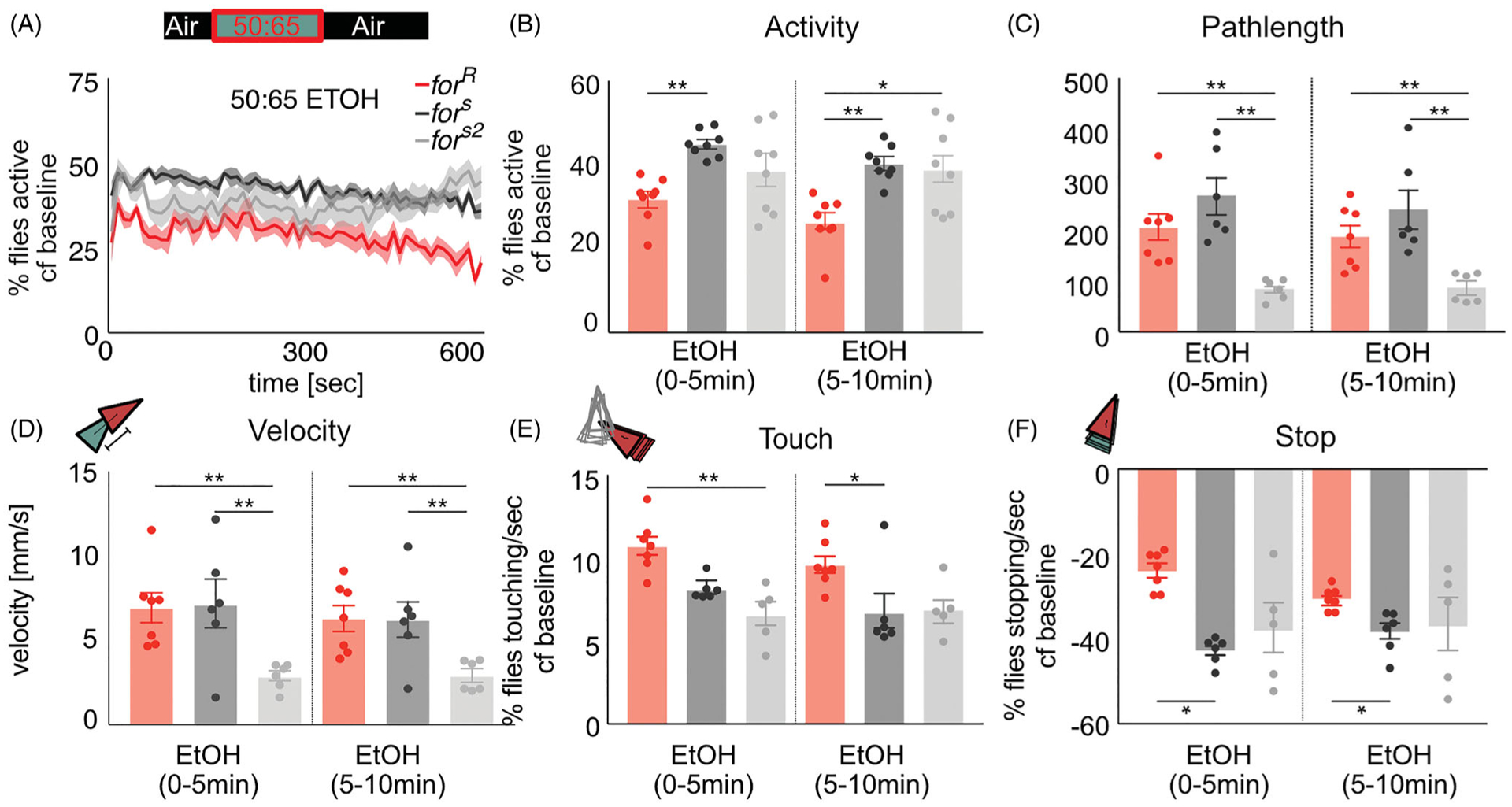
Similar articles
-
Natural polymorphism in protein kinase G modulates functional senescence in Drosophilamelanogaster.J Exp Biol. 2019 Apr 9;222(Pt 7):jeb199364. doi: 10.1242/jeb.199364. J Exp Biol. 2019. PMID: 30910834
-
Activity of cGMP-dependent protein kinase (PKG) affects sucrose responsiveness and habituation in Drosophila melanogaster.Learn Mem. 2004 May-Jun;11(3):303-11. doi: 10.1101/lm.71604. Learn Mem. 2004. PMID: 15169860 Free PMC article.
-
Contribution of a natural polymorphism in protein kinase G modulates electroconvulsive seizure recovery in Drosophila melanogaster.J Exp Biol. 2018 Jul 18;221(Pt 14):jeb179747. doi: 10.1242/jeb.179747. J Exp Biol. 2018. PMID: 29798846
-
The foraging Gene and Its Behavioral Effects: Pleiotropy and Plasticity.Annu Rev Genet. 2019 Dec 3;53:373-392. doi: 10.1146/annurev-genet-112618-043536. Epub 2019 Sep 5. Annu Rev Genet. 2019. PMID: 31487469 Review.
-
Studying alcohol use disorder using Drosophila melanogaster in the era of 'Big Data'.Behav Brain Funct. 2019 Apr 16;15(1):7. doi: 10.1186/s12993-019-0159-x. Behav Brain Funct. 2019. PMID: 30992041 Free PMC article. Review.
Cited by
-
Methods for Exploring the Circuit Basis of Ethanol-Induced Changes in Drosophila Group Locomotor Activity.Cold Spring Harb Protoc. 2023 Oct 3;2023(10):108138. doi: 10.1101/pdb.prot108138. Cold Spring Harb Protoc. 2023. PMID: 37019608 Free PMC article.
-
The foraging gene coordinates brain and heart networks to modulate socially cued interval timing in Drosophila.PLoS Genet. 2025 Jul 8;21(7):e1011752. doi: 10.1371/journal.pgen.1011752. eCollection 2025 Jul. PLoS Genet. 2025. PMID: 40627675 Free PMC article.
References
-
- Alwash N, Allen A, Sokolowski MB, & Levine J (2021). The Drosophila melanogaster foraging gene affects social networks. Journal of Neurogenetics. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases