Comparative Genomics of Peroxisome Biogenesis Proteins: Making Sense of the PEX Proteins
- PMID: 34095119
- PMCID: PMC8172628
- DOI: 10.3389/fcell.2021.654163
Comparative Genomics of Peroxisome Biogenesis Proteins: Making Sense of the PEX Proteins
Abstract
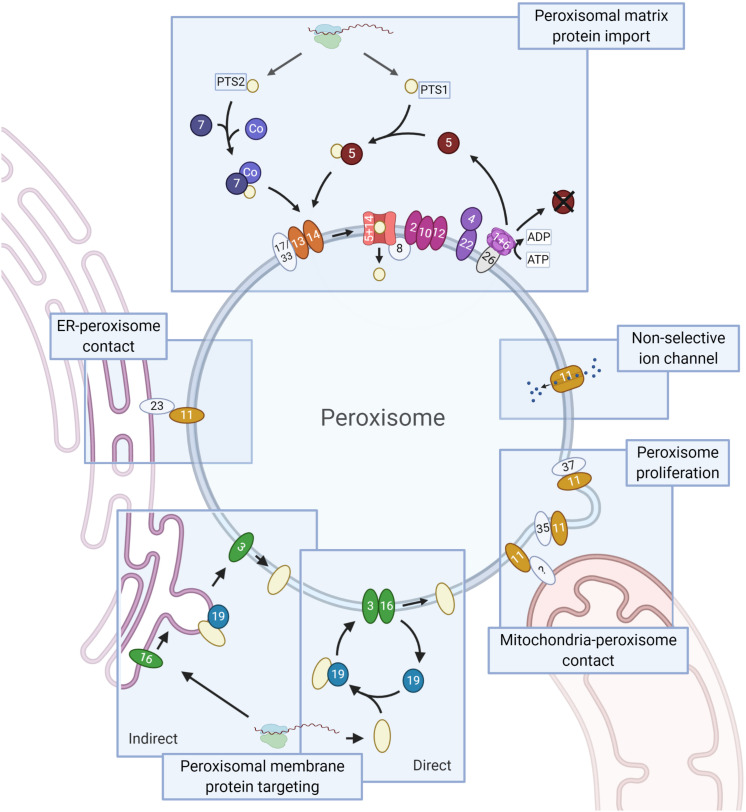
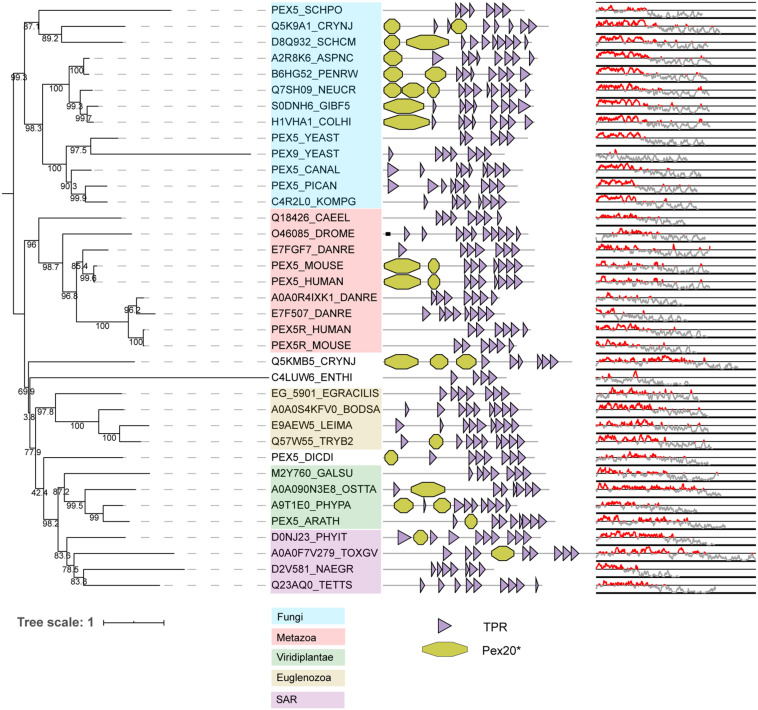
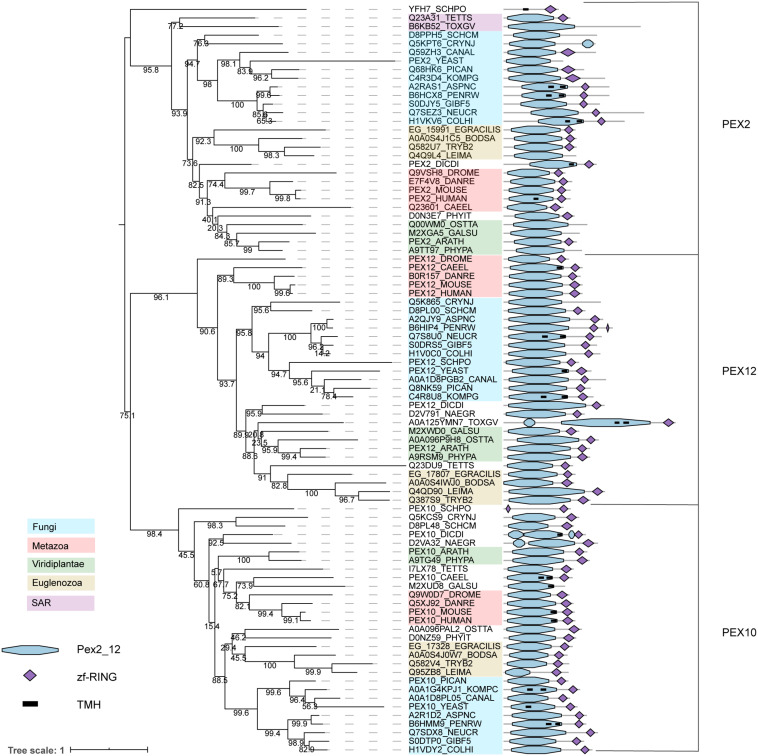
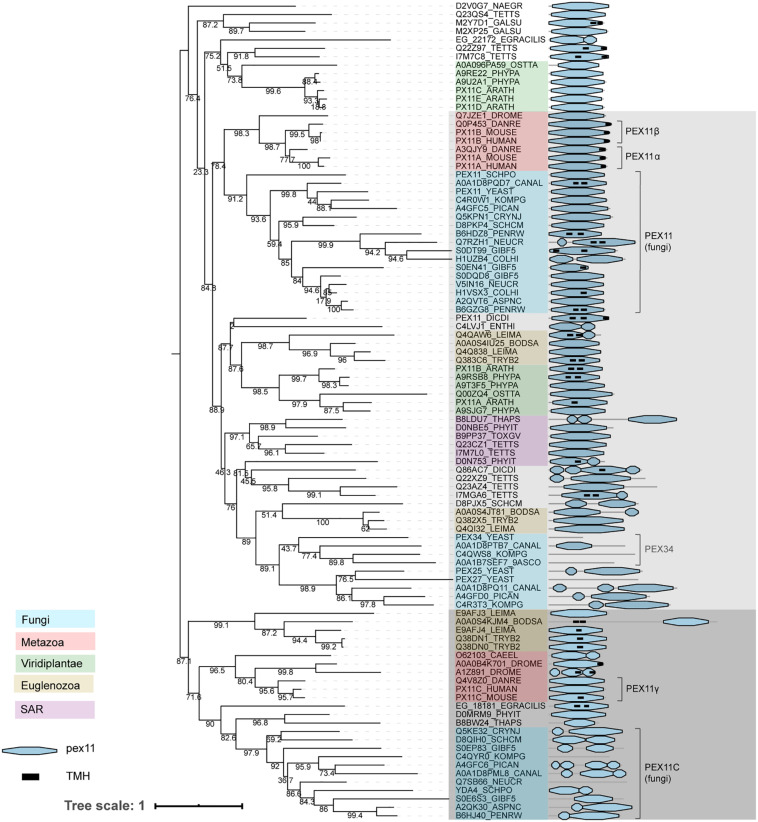
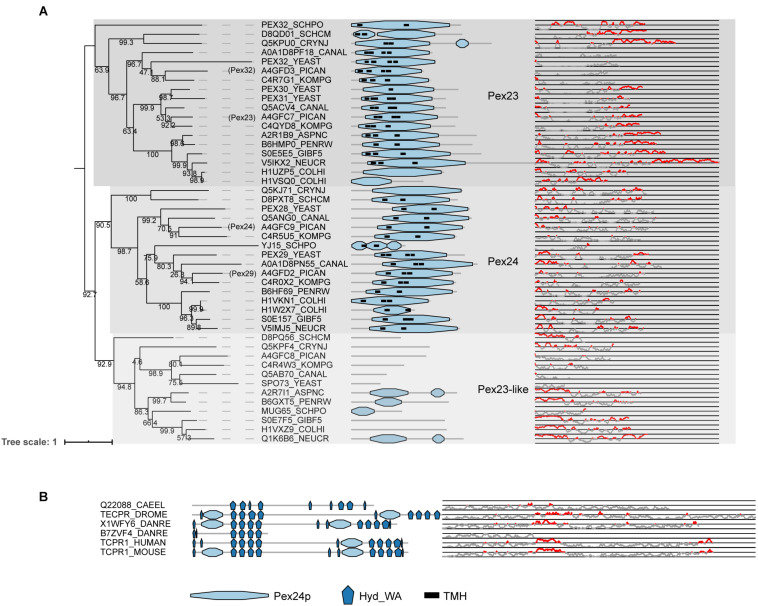
PEX genes encode proteins involved in peroxisome biogenesis and proliferation. Using a comparative genomics approach, we clarify the evolutionary relationships between the 37 known PEX proteins in a representative set of eukaryotes, including all common model organisms, pathogenic unicellular eukaryotes and human. A large number of previously unknown PEX orthologs were identified. We analyzed all PEX proteins, their conservation and domain architecture and defined the core set of PEX proteins that is required to make a peroxisome. The molecular processes in peroxisome biogenesis in different organisms were put into context, showing that peroxisomes are not static organelles in eukaryotic evolution. Organisms that lack peroxisomes still contain a few PEX proteins, which probably play a role in alternative processes. Finally, the relationships between PEX proteins of two large families, the Pex11 and Pex23 families, were analyzed, thereby contributing to the understanding of their complicated and sometimes incorrect nomenclature. We provide an exhaustive overview of this important eukaryotic organelle.
Keywords: PEX; comparative genomics; evolution; peroxisome; protein domains.
Copyright © 2021 Jansen, Santana-Molina, van den Noort, Devos and van der Klei.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures