

The Fkh1 Forkhead associated domain promotes ORC binding to a subset of DNA replication origins in budding yeast
- PMID: 34095951
- PMCID: PMC8501964
- DOI: 10.1093/nar/gkab450
The Fkh1 Forkhead associated domain promotes ORC binding to a subset of DNA replication origins in budding yeast
Abstract
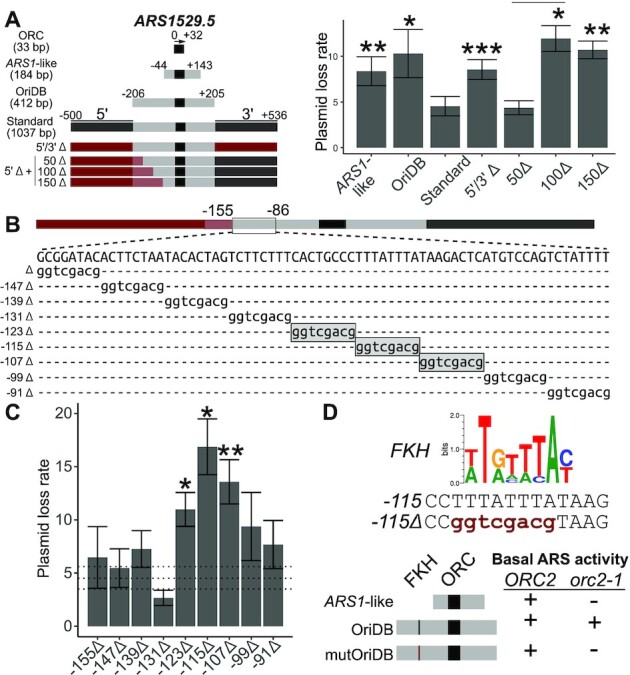
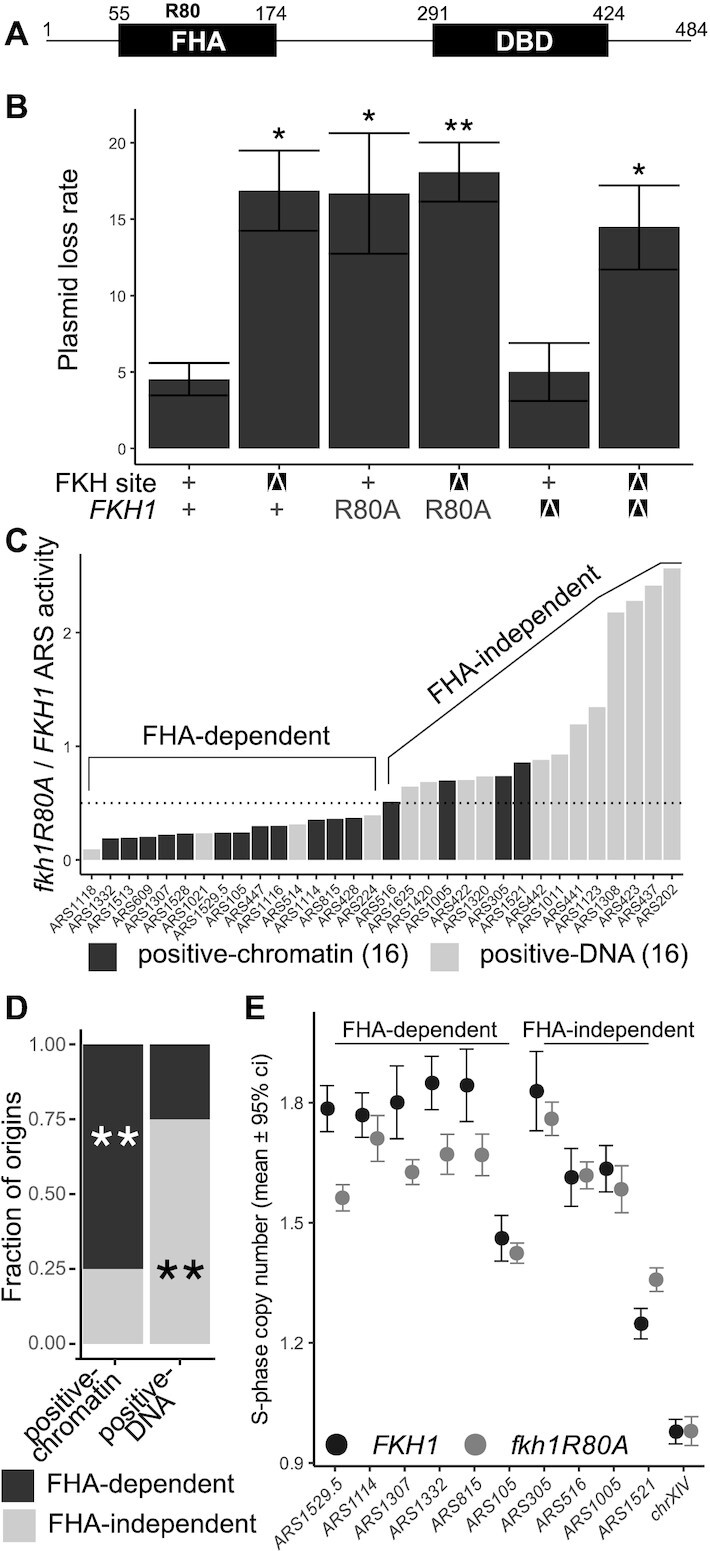
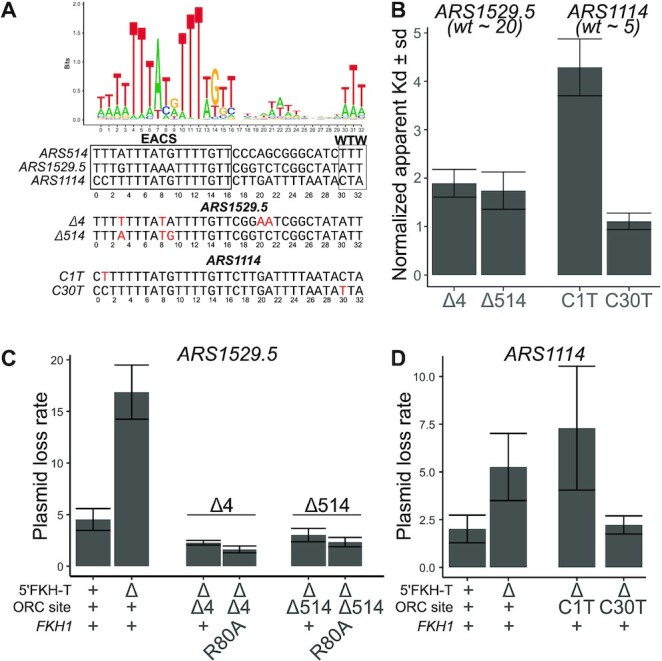
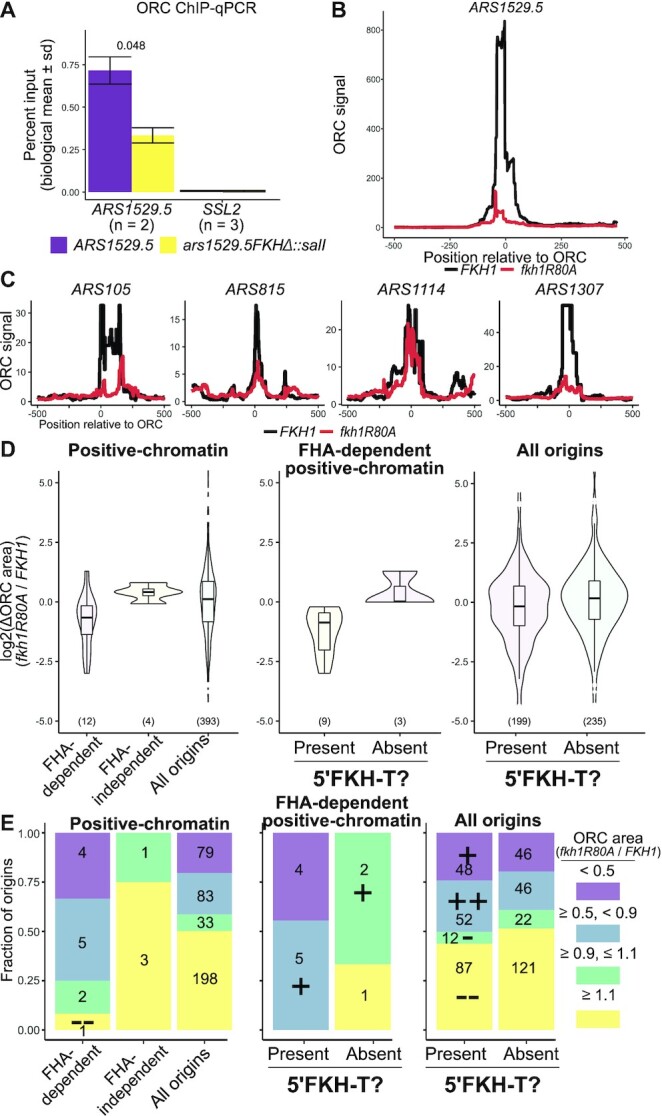
The pioneer event in eukaryotic DNA replication is binding of chromosomal DNA by the origin recognitioncomplex (ORC). The ORC-DNA complex directs the formation of origins, the specific chromosomal regions where DNA synthesis initiates. In all eukaryotes, incompletely understood features of chromatin promote ORC-DNA binding. Here, we uncover a role for the Fkh1 (Forkhead homolog) protein and its forkhead associated (FHA) domain in promoting ORC-origin binding and origin activity at a subset of origins in Saccharomyces cerevisiae. Several of the FHA-dependent origins examined required a distinct Fkh1 binding site located 5' of and proximal to their ORC sites (5'-FKH-T site). Genetic and molecular experiments provided evidence that the Fkh1-FHA domain promoted origin activity directly through Fkh1 binding to this 5' FKH-T site. Nucleotide substitutions within two relevant origins that enhanced their ORC-DNA affinity bypassed the requirement for their 5' FKH-T sites and for the Fkh1-FHA domain. Significantly, assessment of ORC-origin binding by ChIPSeq provided evidence that this mechanism was relevant at ∼25% of yeast origins. Thus, the FHA domain of the conserved cell-cycle transcription factor Fkh1 enhanced origin selection in yeast at the level of ORC-origin binding.
© The Author(s) 2021. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures


Similar articles
-
The budding yeast Fkh1 Forkhead associated (FHA) domain promotes a G1-chromatin state and the activity of chromosomal DNA replication origins.PLoS Genet. 2024 Aug 5;20(8):e1011366. doi: 10.1371/journal.pgen.1011366. eCollection 2024 Aug. PLoS Genet. 2024. PMID: 39102423 Free PMC article.
-
The budding yeast Fkh1 Forkhead associated (FHA) domain promoted a G1-chromatin state and the activity of chromosomal DNA replication origins.bioRxiv [Preprint]. 2024 Jun 7:2024.02.16.580712. doi: 10.1101/2024.02.16.580712. bioRxiv. 2024. Update in: PLoS Genet. 2024 Aug 5;20(8):e1011366. doi: 10.1371/journal.pgen.1011366. PMID: 38405780 Free PMC article. Updated. Preprint.
-
Forkhead transcription factors establish origin timing and long-range clustering in S. cerevisiae.Cell. 2012 Jan 20;148(1-2):99-111. doi: 10.1016/j.cell.2011.12.012. Cell. 2012. PMID: 22265405 Free PMC article.
-
Stepwise assembly of initiation complexes at budding yeast replication origins during the cell cycle.J Cell Sci Suppl. 1995;19:67-72. doi: 10.1242/jcs.1995.supplement_19.9. J Cell Sci Suppl. 1995. PMID: 8655649 Review.
-
Behavior of replication origins in Eukaryota - spatio-temporal dynamics of licensing and firing.Cell Cycle. 2015;14(14):2251-64. doi: 10.1080/15384101.2015.1056421. Epub 2015 Jun 1. Cell Cycle. 2015. PMID: 26030591 Free PMC article. Review.
Cited by
-
Sustained inactivation of the Polycomb PRC1 complex induces DNA repair defects and genomic instability in epigenetic tumors.Histochem Cell Biol. 2024 Jul;162(1-2):133-147. doi: 10.1007/s00418-024-02302-z. Epub 2024 Jun 18. Histochem Cell Biol. 2024. PMID: 38888809 Free PMC article.
-
Gene Expression Analysis of Yeast Strains with a Nonsense Mutation in the eRF3-Coding Gene Highlights Possible Mechanisms of Adaptation.Int J Mol Sci. 2024 Jun 7;25(12):6308. doi: 10.3390/ijms25126308. Int J Mol Sci. 2024. PMID: 38928012 Free PMC article.
-
The budding yeast Fkh1 Forkhead associated (FHA) domain promotes a G1-chromatin state and the activity of chromosomal DNA replication origins.PLoS Genet. 2024 Aug 5;20(8):e1011366. doi: 10.1371/journal.pgen.1011366. eCollection 2024 Aug. PLoS Genet. 2024. PMID: 39102423 Free PMC article.
-
Transcription can be sufficient, but is not necessary, to advance replication timing.bioRxiv [Preprint]. 2025 Feb 5:2025.02.04.636516. doi: 10.1101/2025.02.04.636516. bioRxiv. 2025. PMID: 39975371 Free PMC article. Preprint.
-
Mobile origin-licensing factors confer resistance to conflicts with RNA polymerase.Cell Rep. 2022 Mar 22;38(12):110531. doi: 10.1016/j.celrep.2022.110531. Cell Rep. 2022. PMID: 35320708 Free PMC article.
References
-
- Debatisse M., Le T. B., Letessier A., Dutrillaux B., Brison O.. Common fragile sites: mechanisms of instability revisited. Trends Genet. 2012; 28:22–32. - PubMed
-
- van B. A., Buchanan C., Charboneau E., Fangman W., Brewer B.. An origin-deficient yeast artificial chromosome triggers a cell cycle checkpoint. Mol. Cell. 2001; 7:705–713. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
