

The fibroblast: An emerging key player in thymic T cell selection
- PMID: 34096078
- PMCID: PMC8362222
- DOI: 10.1111/imr.12985
The fibroblast: An emerging key player in thymic T cell selection
Abstract
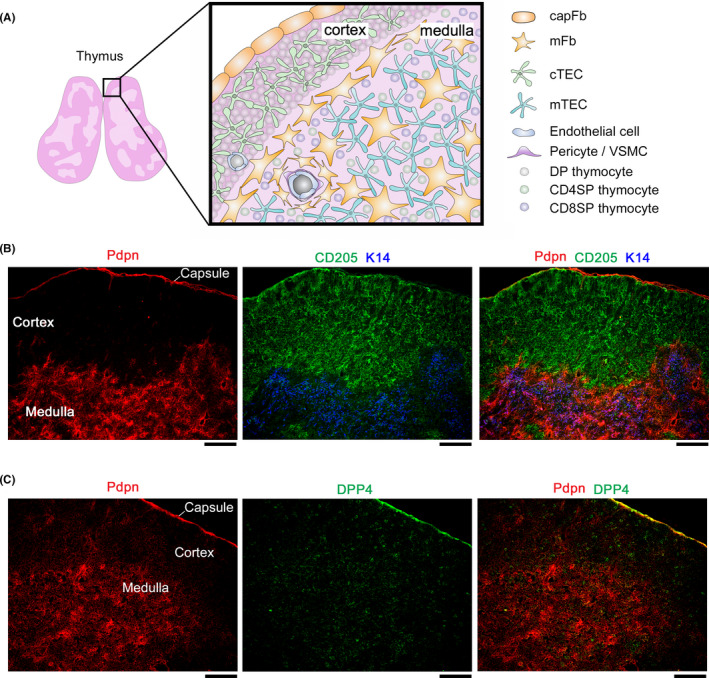
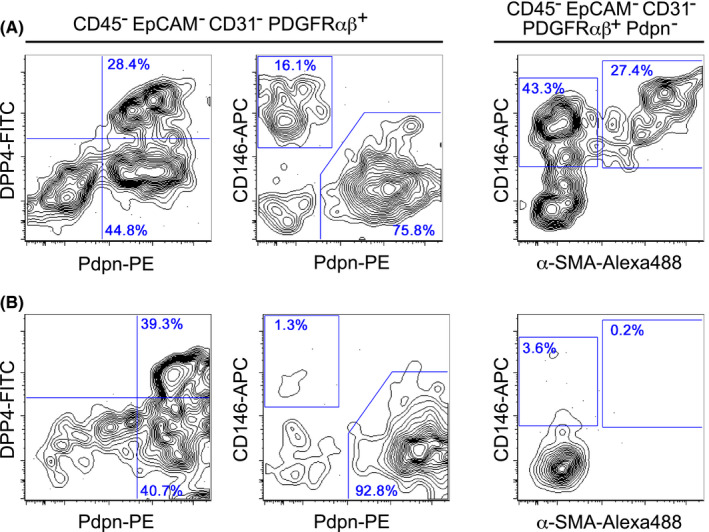
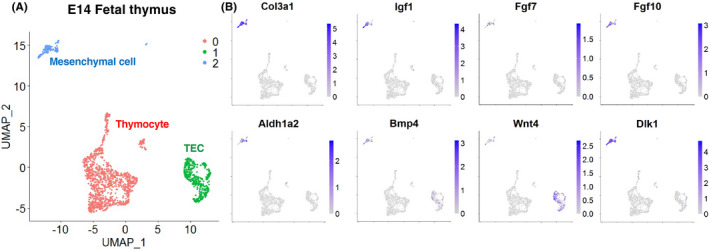
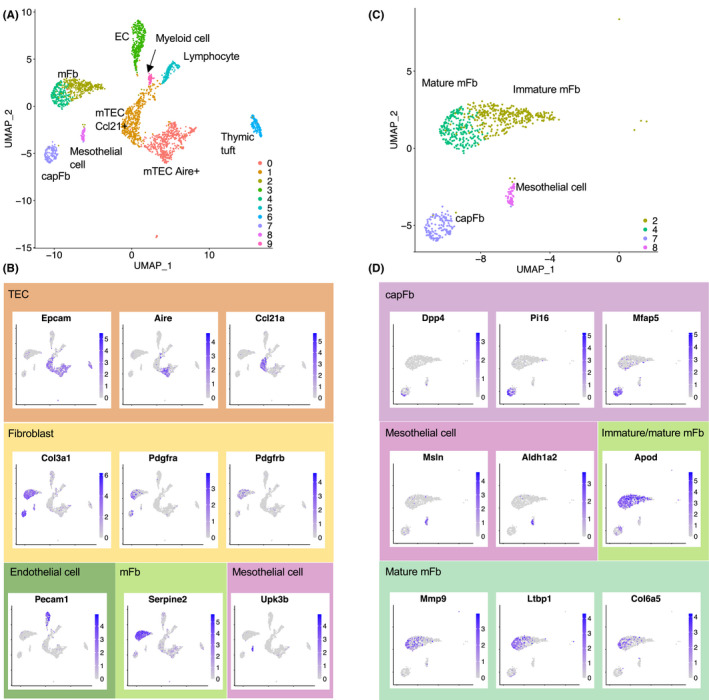
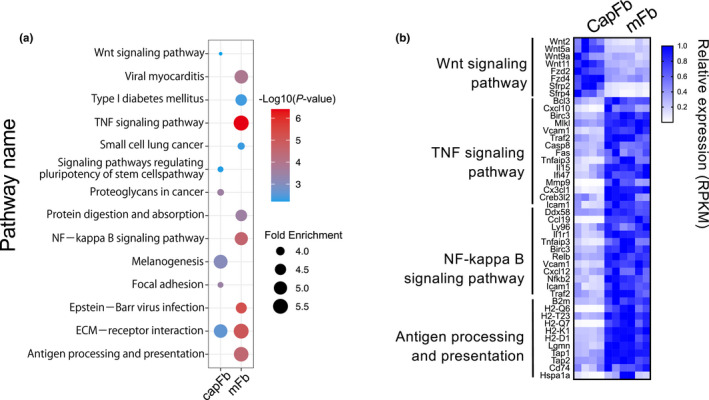
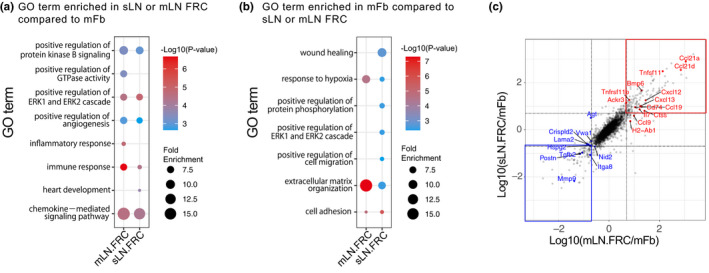
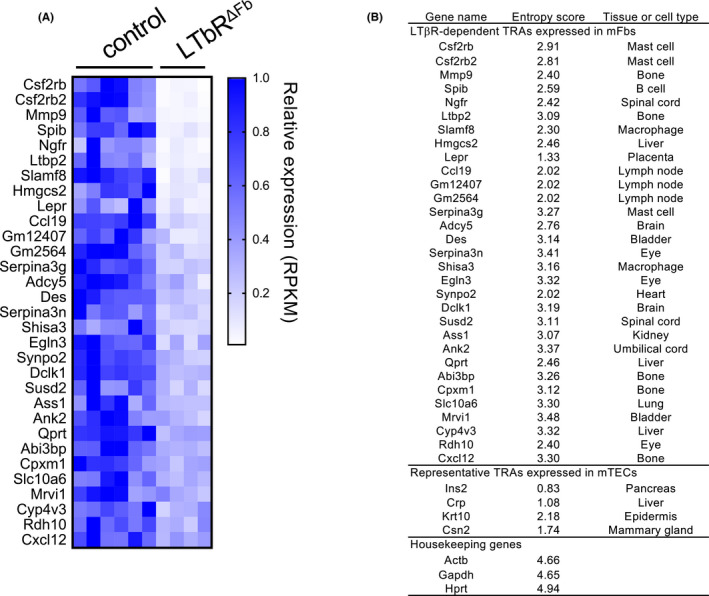
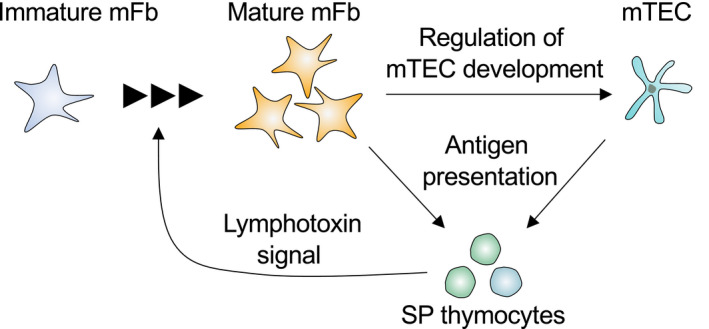
Fibroblasts have recently attracted attention as a key stromal component that controls the immune responses in lymphoid tissues. The thymus has a unique microenvironment comprised of a variety of stromal cells, including fibroblasts and thymic epithelial cells (TECs), the latter of which is known to be important for T cell development because of their ability to express self-antigens. Thymic fibroblasts contribute to thymus organogenesis during embryogenesis and form the capsule and medullary reticular network in the adult thymus. However, the immunological significance of thymic fibroblasts has thus far only been poorly elucidated. In this review, we will summarize the current views on the development and functions of thymic fibroblasts as revealed by new technologies such as multicolor flow cytometry and single cell-based transcriptome profiling. Furthermore, the recently discovered role of medullary fibroblasts in the establishment of T cell tolerance by producing a unique set of self-antigens will be highlighted.
Keywords: T cell; Thymus; capsule; fibroblast; medulla.
© 2021 The Authors. Immunological Reviews published by John Wiley & Sons Ltd.
Conflict of interest statement
There is no conflict of interest to declare.
Figures
References
-
- Miller JFAP. The function of the thymus and its impact on modern medicine. Science. 2020;369(6503):eaba2429. - PubMed
-
- Abramson J, Anderson G. Thymic epithelial cells. Annu Rev Immunol. 2017;35:85‐118. - PubMed
-
- Kadouri N, Nevo S, Goldfarb Y, Abramson J. Thymic epithelial cell heterogeneity: TEC by TEC. Nat Rev Immunol. 2020;20:239‐253. - PubMed
-
- Ohigashi I, Kozai M, Takahama Y. Development and developmental potential of cortical thymic epithelial cells. Immunol Rev. 2016;271:10‐22. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
