

Developmental Exposure to Domoic Acid Disrupts Startle Response Behavior and Circuitry in Zebrafish
- PMID: 34097058
- PMCID: PMC8331133
- DOI: 10.1093/toxsci/kfab066
Developmental Exposure to Domoic Acid Disrupts Startle Response Behavior and Circuitry in Zebrafish
Abstract
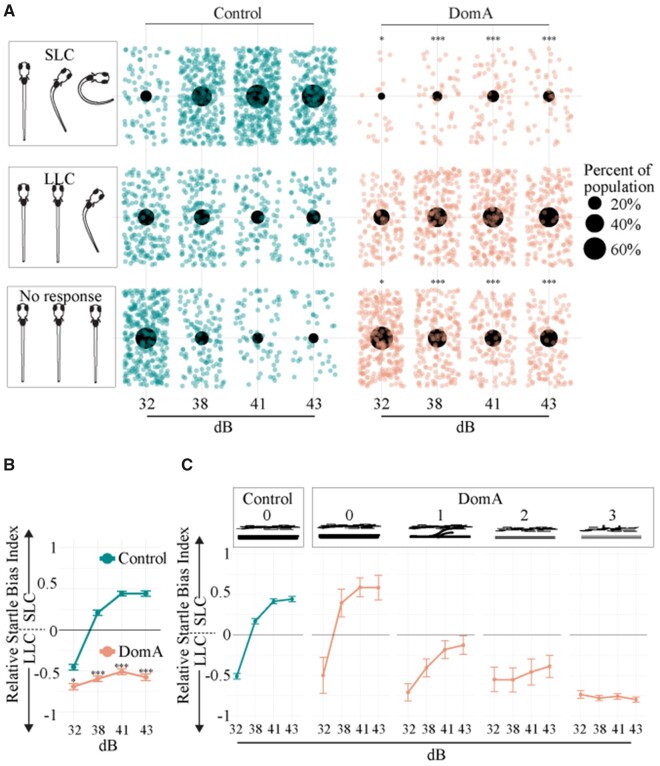
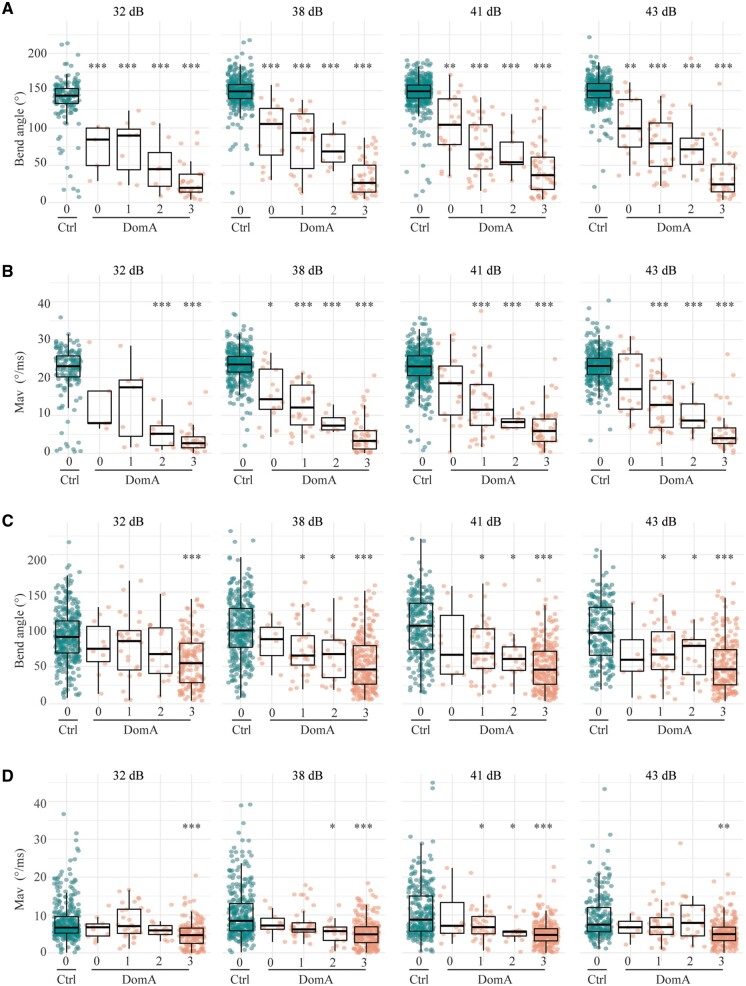
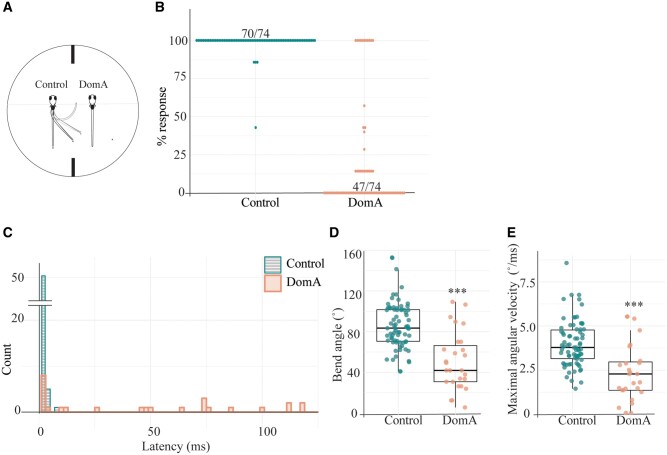
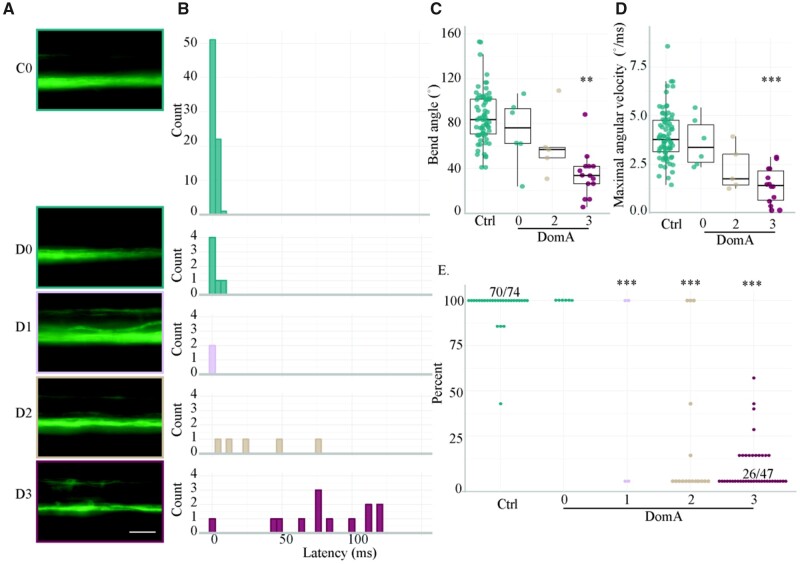
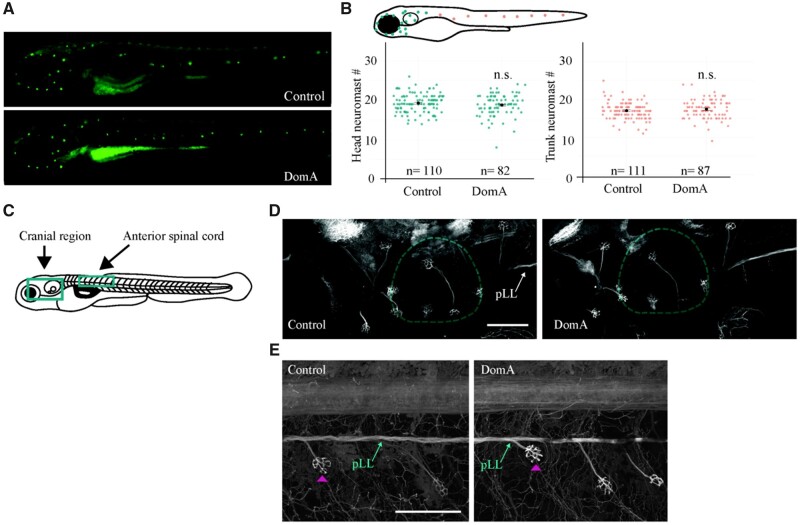
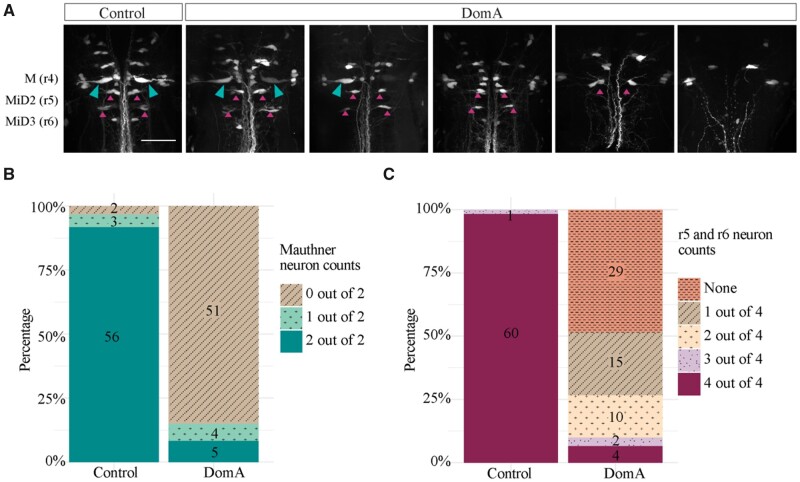
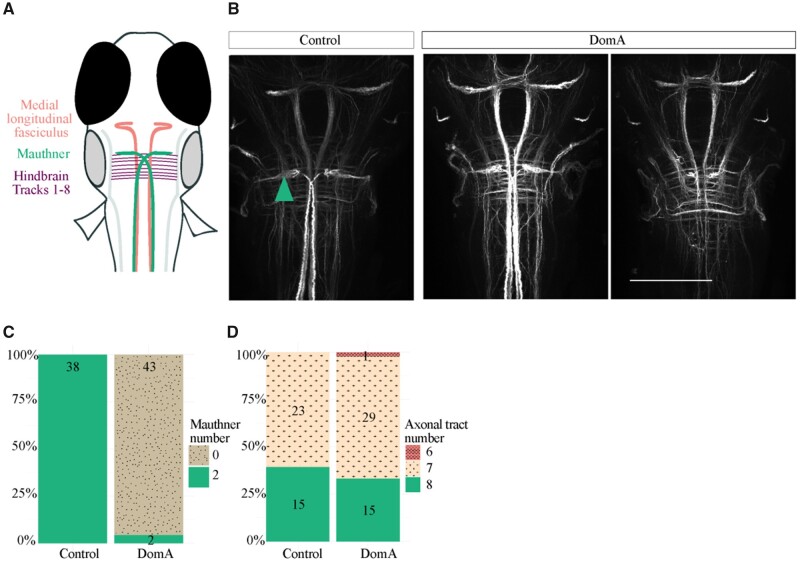
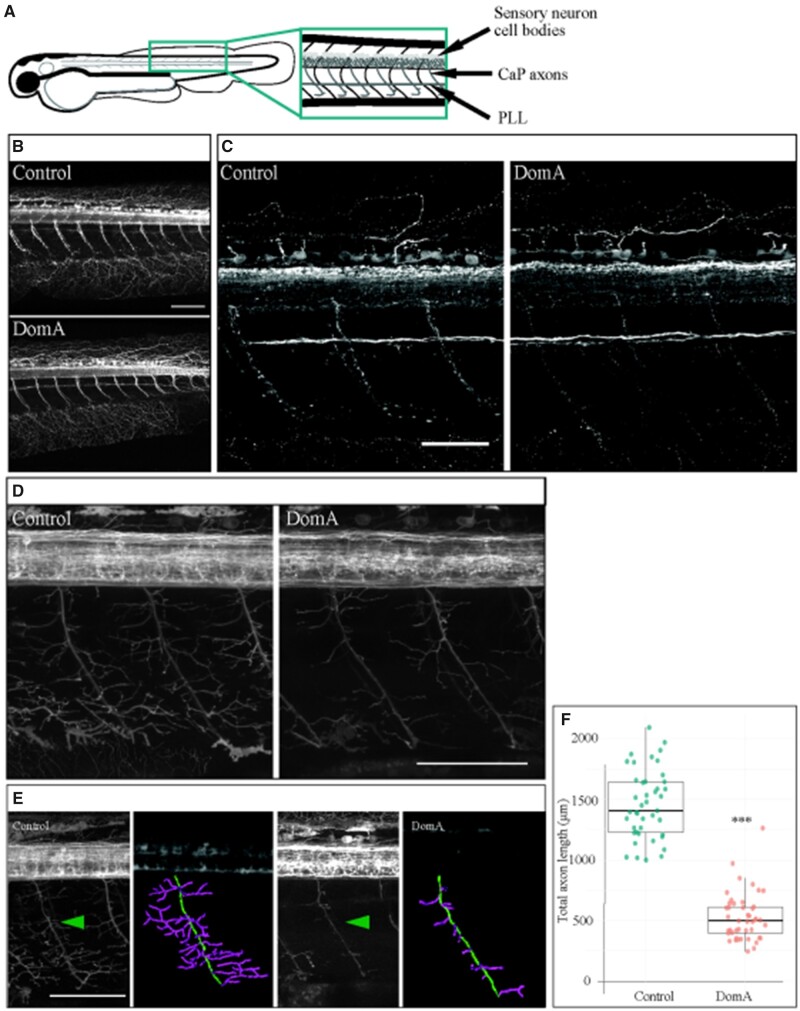
Harmful algal blooms produce potent neurotoxins that accumulate in seafood and are hazardous to human health. Developmental exposure to the harmful algal bloom toxin, domoic acid (DomA), has behavioral consequences well into adulthood, but the cellular and molecular mechanisms of DomA developmental neurotoxicity are largely unknown. To assess these, we exposed zebrafish embryos to DomA during the previously identified window of susceptibility and used the well-known startle response circuit as a tool to identify specific neuronal components that are targeted by exposure to DomA. Exposure to DomA reduced startle responsiveness to both auditory/vibrational and electrical stimuli, and even at the highest stimulus intensities tested, led to a dramatic reduction of one type of startle (short-latency c-starts). Furthermore, DomA-exposed larvae had altered kinematics for both types of startle responses tested, exhibiting shallower bend angles and slower maximal angular velocities. Using vital dye staining, immunolabeling, and live imaging of transgenic lines, we determined that although the sensory inputs were intact, the reticulospinal neurons required for short-latency c-starts were absent in most DomA-exposed larvae. Furthermore, axon tracing revealed that DomA-treated larvae also showed significantly reduced primary motor neuron axon collaterals. Overall, these results show that developmental exposure to DomA targets large reticulospinal neurons and motor neuron axon collaterals, resulting in measurable deficits in startle behavior. They further provide a framework for using the startle response circuit to identify specific neural populations disrupted by toxins or toxicants and to link these disruptions to functional consequences for neural circuit function and behavior.
Keywords: developmental toxicity; domoic acid; escape response; harmful algal bloom toxins; harmful algal blooms; startle circuit; startle response.
© The Author(s) 2021. Published by Oxford University Press on behalf of the Society of Toxicology. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Figures
Similar articles
-
Developmental exposure to domoic acid targets reticulospinal neurons and leads to aberrant myelination in the spinal cord.Sci Rep. 2023 Feb 14;13(1):2587. doi: 10.1038/s41598-023-28166-2. Sci Rep. 2023. PMID: 36788234 Free PMC article.
-
Domoic acid induces developmental stage-specific effects on microglia in zebrafish.Harmful Algae. 2025 Jun;146:102862. doi: 10.1016/j.hal.2025.102862. Epub 2025 Apr 17. Harmful Algae. 2025. PMID: 40409865
-
Developmental Neurotoxicity of the Harmful Algal Bloom Toxin Domoic Acid: Cellular and Molecular Mechanisms Underlying Altered Behavior in the Zebrafish Model.Environ Health Perspect. 2020 Nov;128(11):117002. doi: 10.1289/EHP6652. Epub 2020 Nov 4. Environ Health Perspect. 2020. PMID: 33147070 Free PMC article.
-
Domoic acid as a developmental neurotoxin.Neurotoxicology. 2010 Sep;31(5):409-23. doi: 10.1016/j.neuro.2010.05.003. Epub 2010 May 13. Neurotoxicology. 2010. PMID: 20471419 Free PMC article. Review.
-
Domoic acid and human exposure risks: a review.Toxicon. 2010 Aug 15;56(2):218-30. doi: 10.1016/j.toxicon.2009.05.034. Epub 2009 Jun 6. Toxicon. 2010. PMID: 19505488 Review.
Cited by
-
Developmental exposure of zebrafish to saxitoxin causes altered expression of genes associated with axonal growth.Neurotoxicology. 2024 Dec;105:303-312. doi: 10.1016/j.neuro.2024.11.003. Epub 2024 Nov 19. Neurotoxicology. 2024. PMID: 39571800
-
Developmental exposure to domoic acid targets reticulospinal neurons and leads to aberrant myelination in the spinal cord.Sci Rep. 2023 Feb 14;13(1):2587. doi: 10.1038/s41598-023-28166-2. Sci Rep. 2023. PMID: 36788234 Free PMC article.
-
Domoic acid induces developmental stage-specific effects on microglia in zebrafish.Harmful Algae. 2025 Jun;146:102862. doi: 10.1016/j.hal.2025.102862. Epub 2025 Apr 17. Harmful Algae. 2025. PMID: 40409865
-
Orphan cytochrome P450 20a1 CRISPR/Cas9 mutants and neurobehavioral phenotypes in zebrafish.Sci Rep. 2021 Dec 13;11(1):23892. doi: 10.1038/s41598-021-03068-3. Sci Rep. 2021. PMID: 34903767 Free PMC article.
-
Advancing statistical treatment of photolocomotor behavioral response study data.PLoS One. 2024 May 21;19(5):e0300636. doi: 10.1371/journal.pone.0300636. eCollection 2024. PLoS One. 2024. PMID: 38771799 Free PMC article.
References
-
- Anderson D. M., Glibert P. M., Burkholder J. M. (2002). Harmful algal blooms and eutrophication: Nutrient sources, composition, and consequences. Estuaries 25, 704–726.
-
- Bates, D., Mächler, M., Bolker, B., and Walker, S. (2015). Fitting Linear Mixed-Effects Models using lme4. J. Stat. Softw. 67, 1–48.
-
- Bates S. S., Hubbard K. A., Lundholm N., Montresor M., Leaw C. P. (2018). Pseudo-nitzschia, Nitzschia, and domoic acid: New research since 2011. Harmful Algae 79, 3–43. - PubMed
-
- Benaglia T., Chauveau, D., Hunter, D., and Young, D. (2009). mixtools: An R package for analyzing finite mixture models. J. Stat. Softw. 32, 1–29.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
