

Dopamine, vocalization, and astrocytes
- PMID: 34098250
- PMCID: PMC8260450
- DOI: 10.1016/j.bandl.2021.104970
Dopamine, vocalization, and astrocytes
Abstract
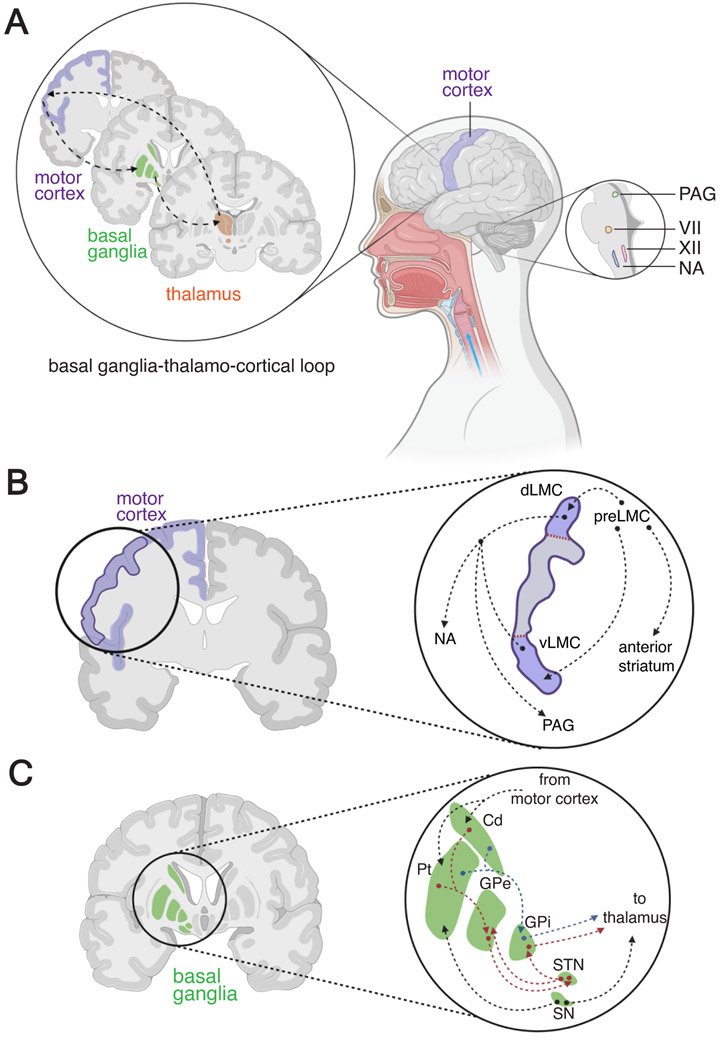
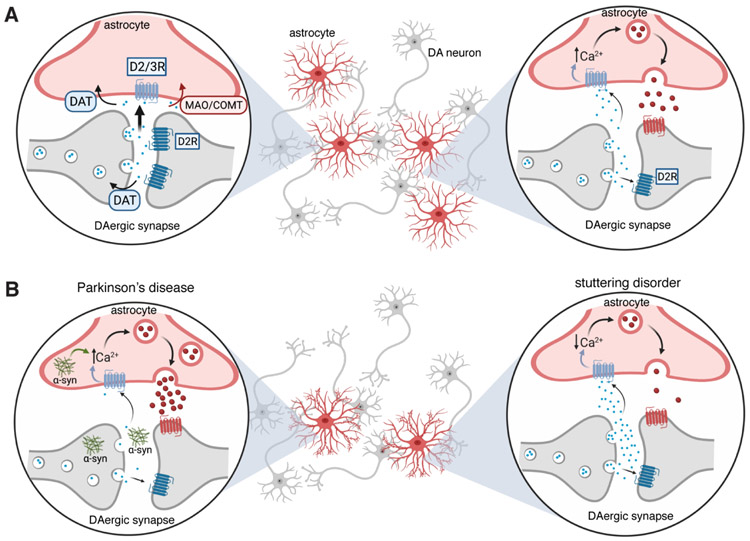
Dopamine, the main catecholamine neurotransmitter in the brain, is predominately produced in the basal ganglia and released to various brain regions including the frontal cortex, midbrain and brainstem. Dopamine's effects are widespread and include modulation of a number of voluntary and innate behaviors. Vigilant regulation and modulation of dopamine levels throughout the brain is imperative for proper execution of motor behaviors, in particular speech and other types of vocalizations. While dopamine's role in motor circuitry is widely accepted, its unique function in normal and abnormal speech production is not fully understood. In this perspective, we first review the role of dopaminergic circuits in vocal production. We then discuss and propose the conceivable involvement of astrocytes, the numerous star-shaped glia cells of the brain, in the dopaminergic network modulating normal and abnormal vocal productions.
Keywords: Astrocytes; Basal ganglia; Dopamine; Glia; Speech; Vocalization.
Copyright © 2021. Published by Elsevier Inc.
Conflict of interest statement
Figures
Similar articles
-
Morphometric analysis of astrocytes in vocal production circuits of common marmoset (Callithrix jacchus).J Comp Neurol. 2022 Feb;530(2):574-589. doi: 10.1002/cne.25230. Epub 2021 Aug 23. J Comp Neurol. 2022. PMID: 34387357 Free PMC article.
-
Social modulation of learned behavior by dopamine in the basal ganglia: insights from songbirds.J Physiol Paris. 2013 Jun;107(3):219-29. doi: 10.1016/j.jphysparis.2012.09.002. Epub 2012 Sep 29. J Physiol Paris. 2013. PMID: 23032272 Review.
-
Dysbindin-1A modulation of astrocytic dopamine and basal ganglia dependent behaviors relevant to schizophrenia.Mol Psychiatry. 2022 Oct;27(10):4201-4217. doi: 10.1038/s41380-022-01683-8. Epub 2022 Jul 11. Mol Psychiatry. 2022. PMID: 35821415
-
Astrocytic modulation of central pattern generating motor circuits.Glia. 2022 Aug;70(8):1506-1519. doi: 10.1002/glia.24162. Epub 2022 Feb 25. Glia. 2022. PMID: 35212422 Review.
-
Modulating the map: dopaminergic tuning of hippocampal spatial coding and interactions.Prog Brain Res. 2015;219:187-216. doi: 10.1016/bs.pbr.2015.03.002. Epub 2015 May 16. Prog Brain Res. 2015. PMID: 26072240 Review.
Cited by
-
Neurobiology of Obsessive-Compulsive Disorder from Genes to Circuits: Insights from Animal Models.Neurosci Bull. 2024 Dec;40(12):1975-1994. doi: 10.1007/s12264-024-01252-9. Epub 2024 Jul 9. Neurosci Bull. 2024. PMID: 38982026 Review.
-
Assessing Different Histological Preparations for Reconstruction of Astrocyte Tridimensional Structure.Cells. 2024 Jun 3;13(11):969. doi: 10.3390/cells13110969. Cells. 2024. PMID: 38891101 Free PMC article.
-
Neuroscience of cancer: unraveling the complex interplay between the nervous system, the tumor and the tumor immune microenvironment.Mol Cancer. 2025 Jan 17;24(1):24. doi: 10.1186/s12943-024-02219-0. Mol Cancer. 2025. PMID: 39825376 Free PMC article. Review.
-
Repurposed Antipsychotics as Potential Anticancer Agents: Clozapine Efficacy and Dopaminergic Pathways in Neuroblastoma and Glioblastoma.Life (Basel). 2025 Jul 12;15(7):1097. doi: 10.3390/life15071097. Life (Basel). 2025. PMID: 40724598 Free PMC article.
-
Probing midbrain dopamine function in pediatric obsessive-compulsive disorder via neuromelanin-sensitive magnetic resonance imaging.Mol Psychiatry. 2023 Jul;28(7):3075-3082. doi: 10.1038/s41380-023-02105-z. Epub 2023 May 17. Mol Psychiatry. 2023. PMID: 37198261 Free PMC article.
References
-
- Abdalla A, West A, Jin Y, Saylor RA, Qiang B, Peña E, … Hashemi P (2020). Fast serotonin voltammetry as a versatile tool for mapping dynamic tissue architecture: I. Responses at carbon fibers describe local tissue physiology. Journal of Neurochemistry, 153(1), 33–50. doi:10.1111/jnc.14854 - DOI - PMC - PubMed
-
- American Psychiatric Association. (2013). Diagnostic And Statistical Manual Of Mental Disorders, 5th Edition: Dsm-5 (5th ed., p. 991). Washington, D.C: American Psychiatric Publishing.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
