

Effects of short-chain fatty acids in inhibiting HDAC and activating p38 MAPK are critical for promoting B10 cell generation and function
- PMID: 34099635
- PMCID: PMC8184914
- DOI: 10.1038/s41419-021-03880-9
Effects of short-chain fatty acids in inhibiting HDAC and activating p38 MAPK are critical for promoting B10 cell generation and function
Abstract
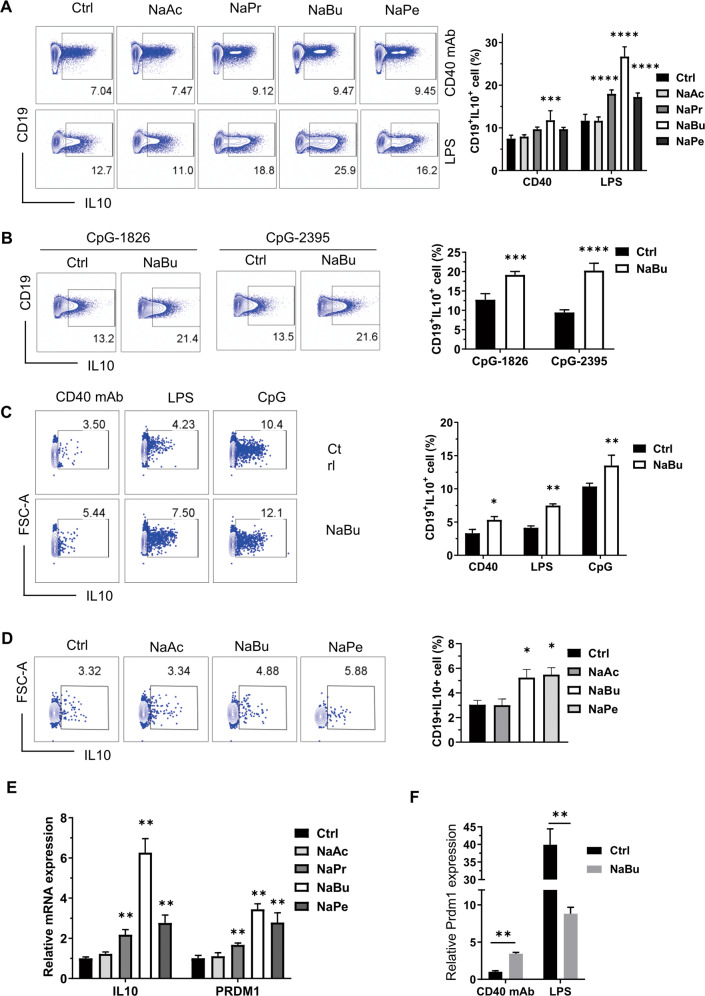
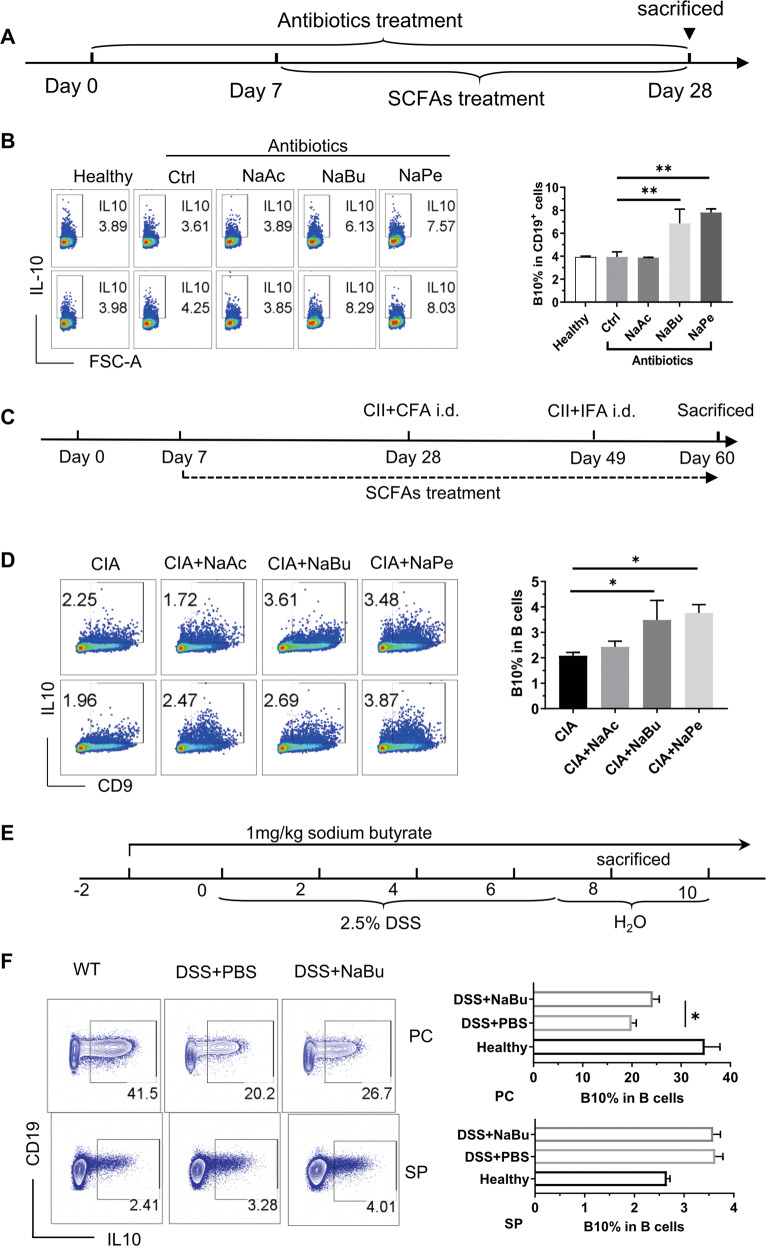
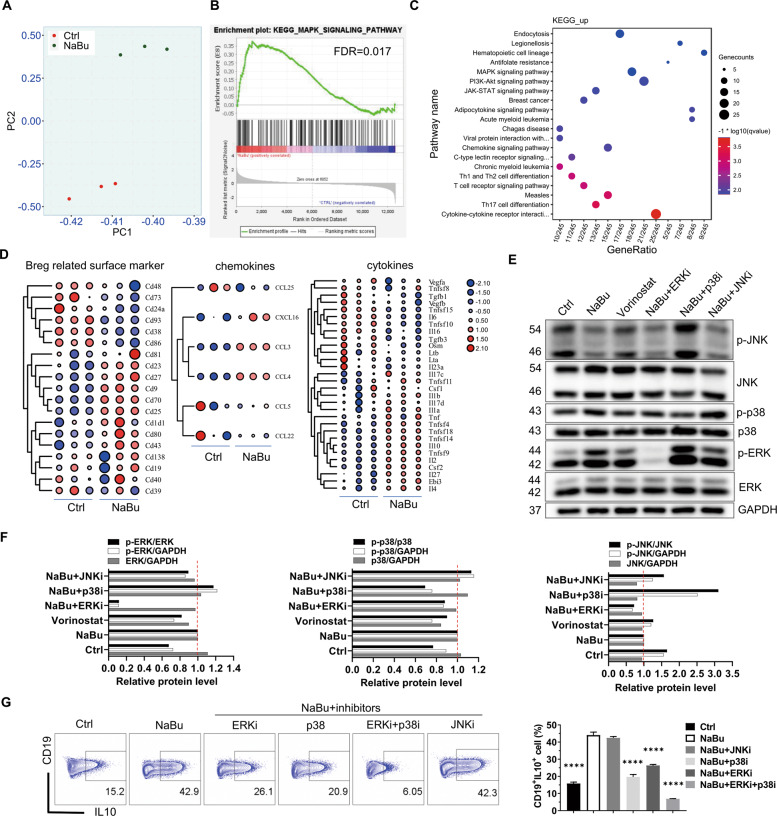
B10 cells are regulatory B cells capable of producing IL-10 for maintaining immune homeostasis. Dysregulation of B10 cells occurs in autoimmune and inflammatory diseases. Modulation or adoptive transfer of B10 cells is a promising therapeutic strategy. The short-chain fatty acids (SCFAs), the metabolites of microbiota, play a critical role in maintaining immune homeostasis and are the potential drugs for the modulation of B10 cells. It is not clear whether and how SCFAs upregulate the frequency of B10 cells. Here, we found that SCFAs could promote murine and human B10 cell generation in vitro. Upregulation of B10 cells by butyrate or pentanoate was also observed in either healthy mice, mice with dextran sodium sulfate (DSS)-induced colitis, or mice with collagen-induced arthritis. Moreover, SCFA treatment could ameliorate clinical scores of colitis and arthritis. Adoptive transfer of B cells pretreated with butyrate showed more alleviation of DSS-induced colitis than those without butyrate. A further study demonstrates that SCFAs upregulate B10 cells in a manner dependent on their histone deacetylase (HDAC) inhibitory activity and independent of the G-protein-coupled receptor pathway. Transcriptomic analysis indicated that the MAPK signaling pathway was enriched in B10 cells treated with butyrate. A study with inhibitors of ERK, JNK, and p38 MAPK demonstrated that activating p38 MAPK by butyrate is critical for the upregulation of B10 cells. Moreover, HDAC inhibitor has similar effects on B10 cells. Our study sheds light on the mechanism underlying B10 cell differentiation and function and provides a potential therapeutic strategy with SCFAs and HDAC inhibitors for inflammation and autoimmune diseases.
Conflict of interest statement
The authors declare no competing interests.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
