

Light regulates stomatal development by modulating paracrine signaling from inner tissues
- PMID: 34099707
- PMCID: PMC8184810
- DOI: 10.1038/s41467-021-23728-2
Light regulates stomatal development by modulating paracrine signaling from inner tissues
Abstract
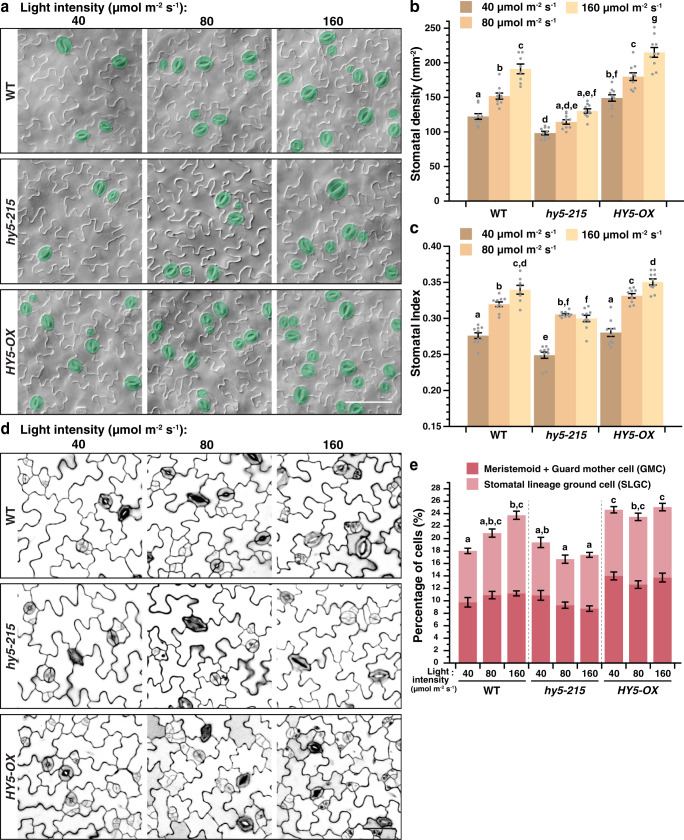
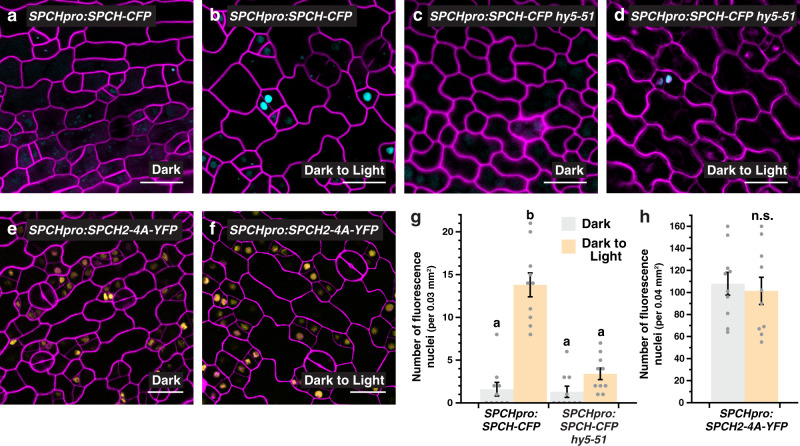
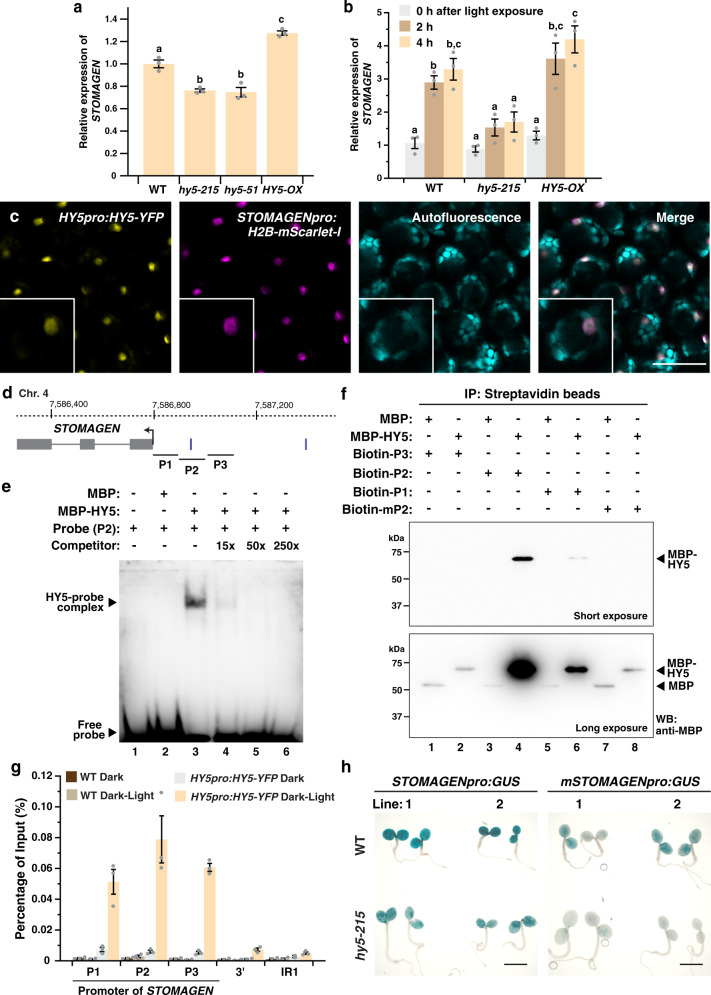
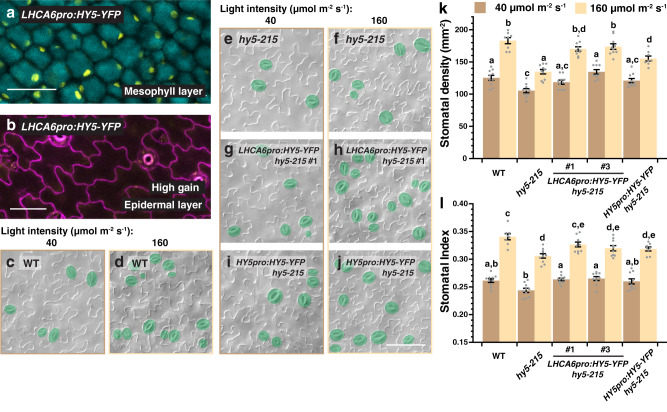
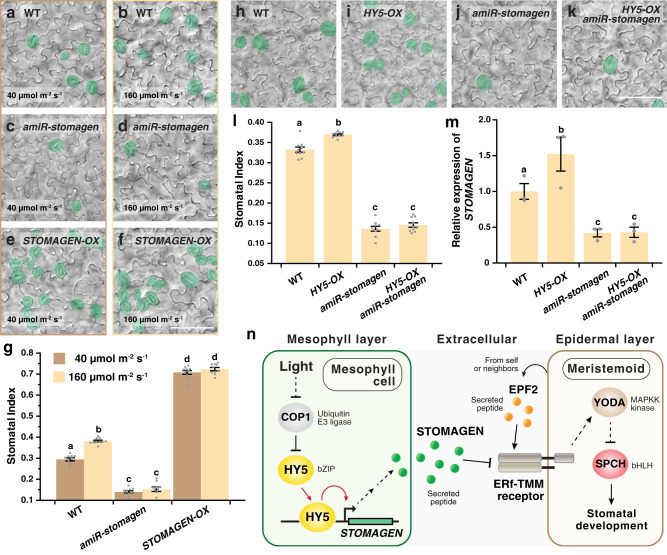
Developmental outcomes are shaped by the interplay between intrinsic and external factors. The production of stomata-essential pores for gas exchange in plants-is extremely plastic and offers an excellent system to study this interplay at the cell lineage level. For plants, light is a key external cue, and it promotes stomatal development and the accumulation of the master stomatal regulator SPEECHLESS (SPCH). However, how light signals are relayed to influence SPCH remains unknown. Here, we show that the light-regulated transcription factor ELONGATED HYPOCOTYL 5 (HY5), a critical regulator for photomorphogenic growth, is present in inner mesophyll cells and directly binds and activates STOMAGEN. STOMAGEN, the mesophyll-derived secreted peptide, in turn stabilizes SPCH in the epidermis, leading to enhanced stomatal production. Our work identifies a molecular link between light signaling and stomatal development that spans two tissue layers and highlights how an environmental signaling factor may coordinate growth across tissue types.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
