

Hippocampal place cell sequences differ during correct and error trials in a spatial memory task
- PMID: 34099727
- PMCID: PMC8185092
- DOI: 10.1038/s41467-021-23765-x
Hippocampal place cell sequences differ during correct and error trials in a spatial memory task
Abstract
Theta rhythms temporally coordinate sequences of hippocampal place cell ensembles during active behaviors, while sharp wave-ripples coordinate place cell sequences during rest. We investigated whether such coordination of hippocampal place cell sequences is disrupted during error trials in a delayed match-to-place task. As a reward location was learned across trials, place cell sequences developed that represented temporally compressed paths to the reward location during the approach to the reward location. Less compressed paths were represented on error trials as an incorrect stop location was approached. During rest periods of correct but not error trials, place cell sequences developed a bias to replay representations of paths ending at the correct reward location. These results support the hypothesis that coordination of place cell sequences by theta rhythms and sharp wave-ripples develops as a reward location is learned and may be important for the successful performance of a spatial memory task.
Conflict of interest statement
The authors declare no competing interests.
Figures
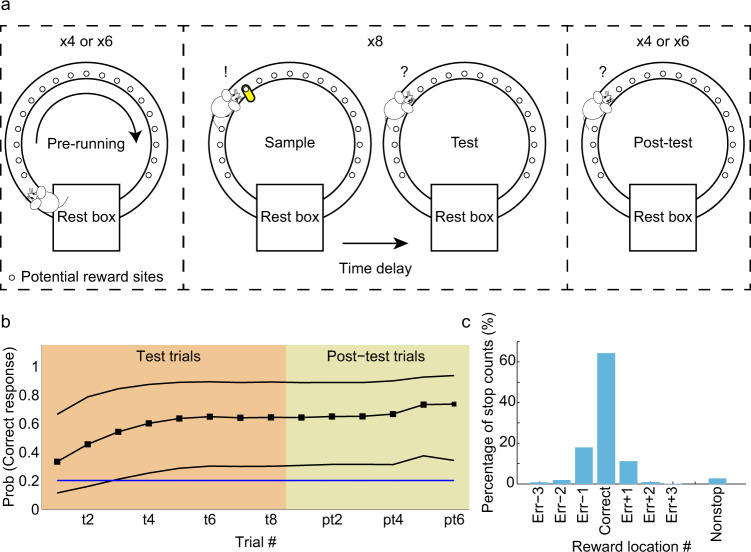

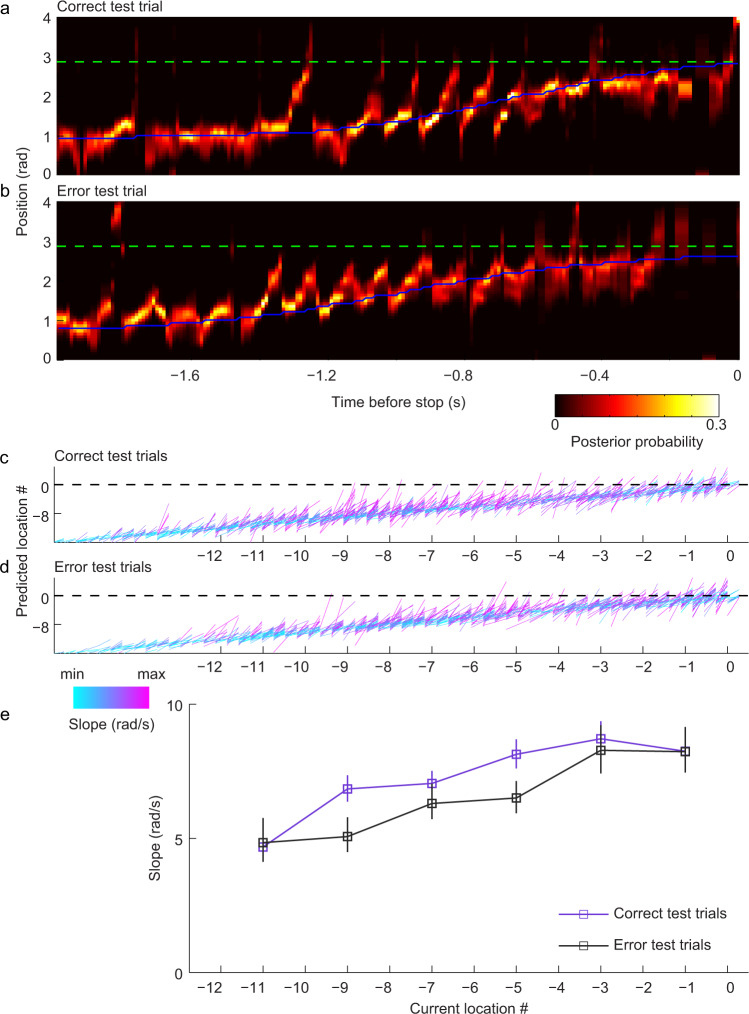
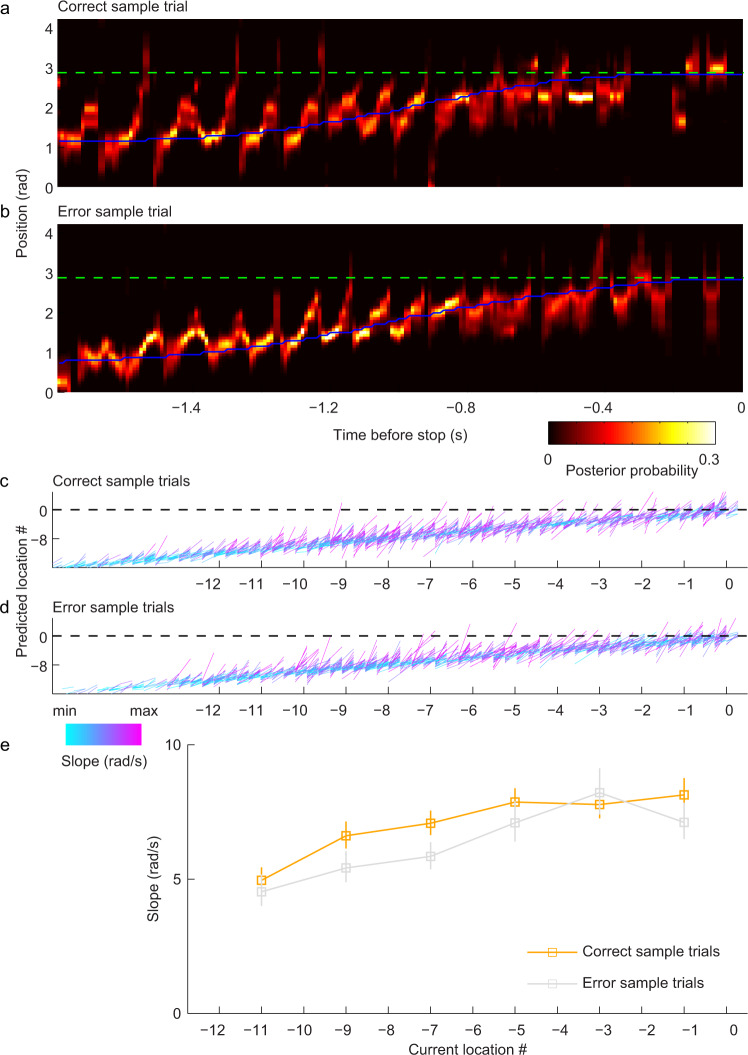
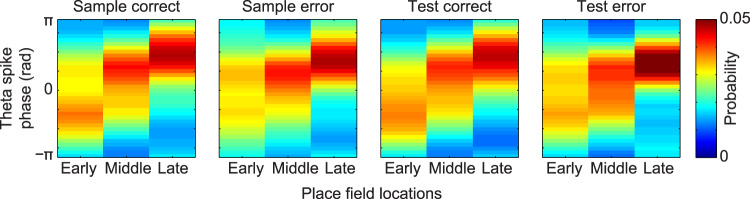



References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
