

Genetically Encoded, pH-Sensitive mTFP1 Biosensor for Probing Lysosomal pH
- PMID: 34102054
- PMCID: PMC8240087
- DOI: 10.1021/acssensors.0c02318
Genetically Encoded, pH-Sensitive mTFP1 Biosensor for Probing Lysosomal pH
Abstract
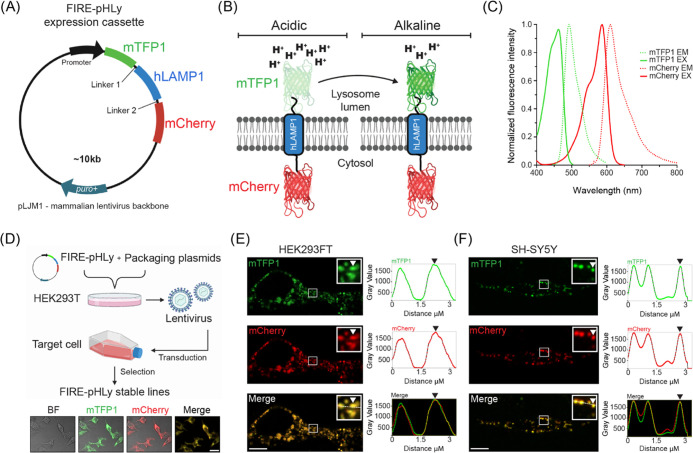
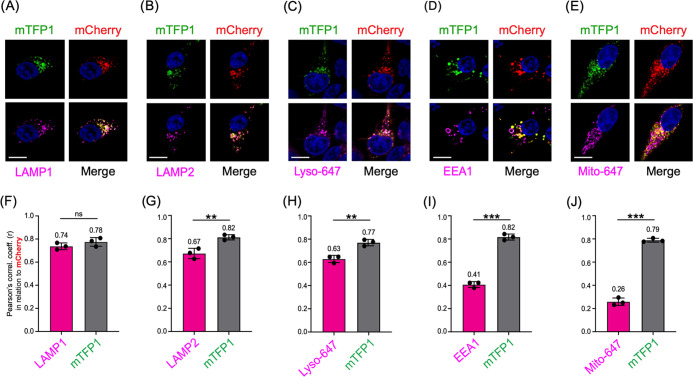
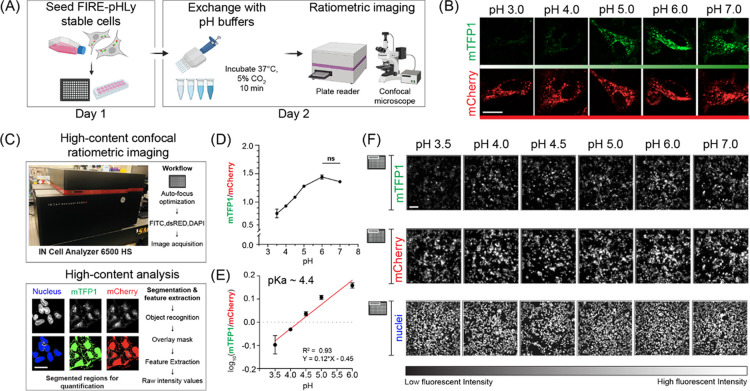
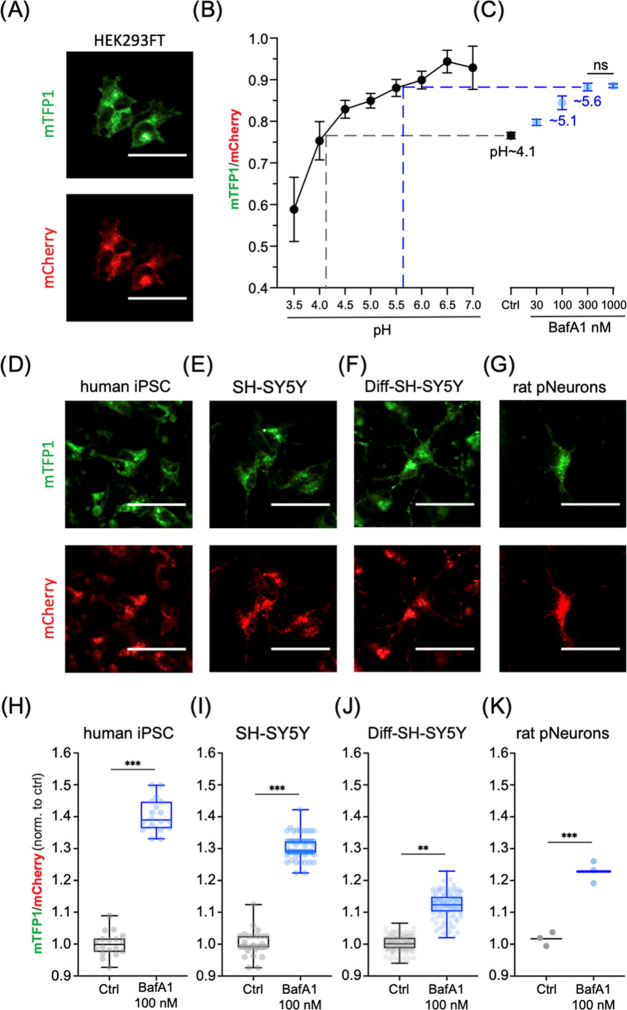
Lysosomes are important sites for macromolecular degradation, defined by an acidic lumenal pH of ∼4.5. To better understand lysosomal pH, we designed a novel, genetically encoded, fluorescent protein (FP)-based pH biosensor called Fluorescence Indicator REporting pH in Lysosomes (FIRE-pHLy). This biosensor was targeted to lysosomes with lysosomal-associated membrane protein 1 (LAMP1) and reported lumenal pH between 3.5 and 6.0 with monomeric teal fluorescent protein 1 (mTFP1), a bright cyan pH-sensitive FP variant with a pKa of 4.3. Ratiometric quantification was enabled with cytosolically oriented mCherry using high-content quantitative imaging. We expressed FIRE-pHLy in several cellular models and quantified the alkalinizing response to bafilomycin A1, a specific V-ATPase inhibitor. In summary, we have engineered FIRE-pHLy, a specific, robust, and versatile lysosomal pH biosensor, that has broad applications for investigating pH dynamics in aging- and lysosome-related diseases, as well as in lysosome-based drug discovery.
Keywords: high-content analysis; lysosomes; neurons; pH biosensor; ratiometric imaging.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
