

A versatile reverse genetics platform for SARS-CoV-2 and other positive-strand RNA viruses
- PMID: 34103499
- PMCID: PMC8187723
- DOI: 10.1038/s41467-021-23779-5
A versatile reverse genetics platform for SARS-CoV-2 and other positive-strand RNA viruses
Abstract
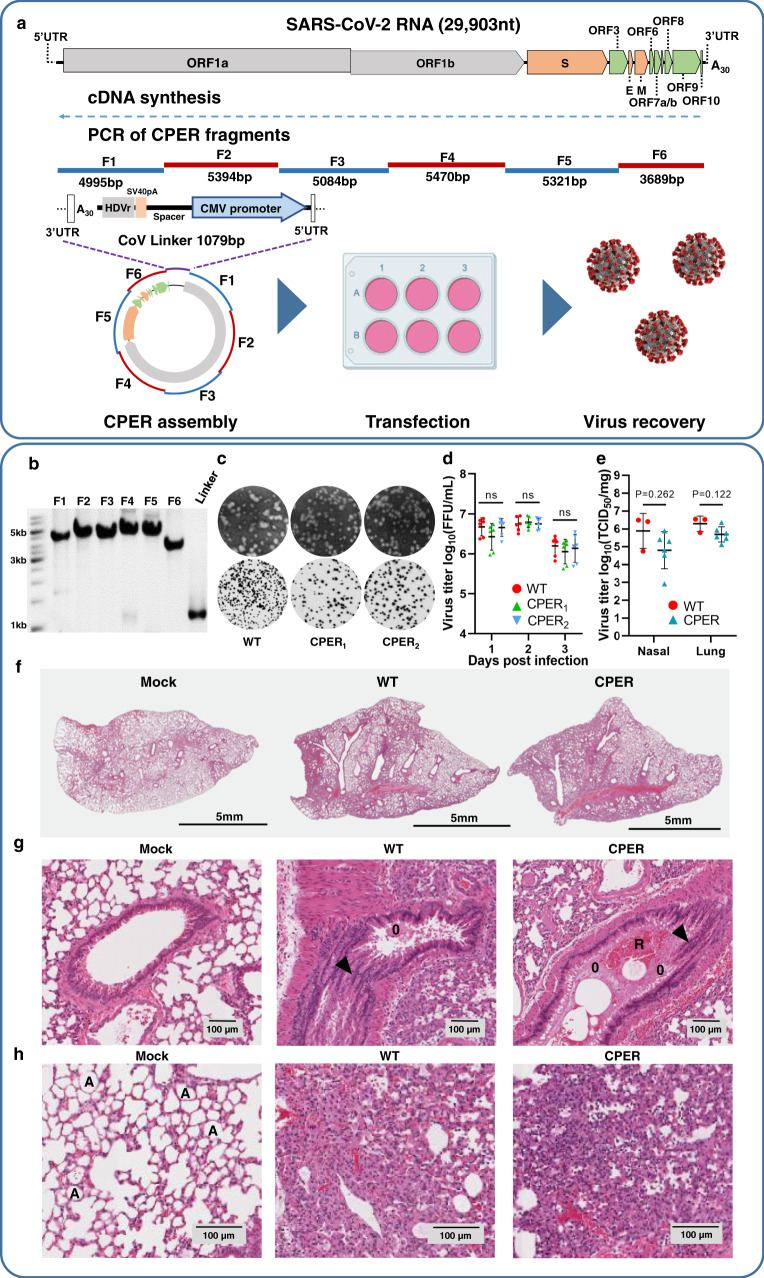
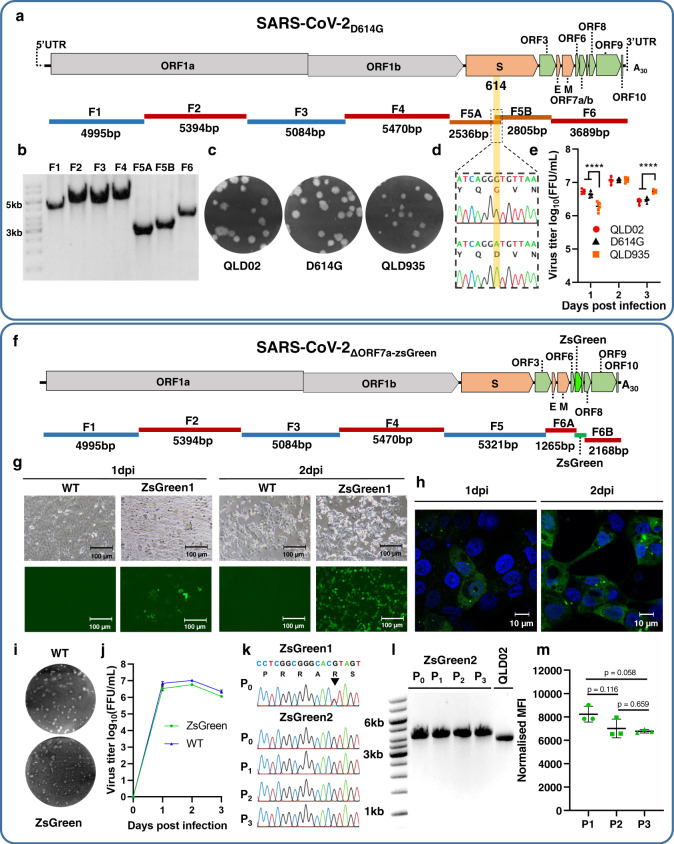
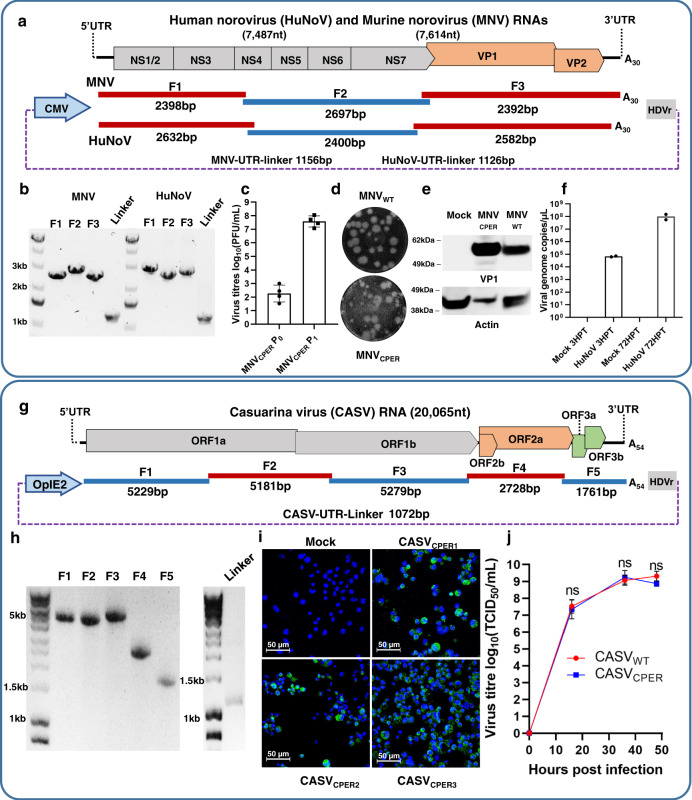
The current COVID-19 pandemic is caused by the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). We demonstrate that despite the large size of the viral RNA genome (~30 kb), infectious full-length cDNA is readily assembled in vitro by a circular polymerase extension reaction (CPER) methodology without the need for technically demanding intermediate steps. Overlapping cDNA fragments are generated from viral RNA and assembled together with a linker fragment containing CMV promoter into a circular full-length viral cDNA in a single reaction. Transfection of the circular cDNA into mammalian cells results in the recovery of infectious SARS-CoV-2 virus that exhibits properties comparable to the parental virus in vitro and in vivo. CPER is also used to generate insect-specific Casuarina virus with ~20 kb genome and the human pathogens Ross River virus (Alphavirus) and Norovirus (Calicivirus), with the latter from a clinical sample. Additionally, reporter and mutant viruses are generated and employed to study virus replication and virus-receptor interactions.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
