

Sleep deprivation results in diverse patterns of synaptic scaling across the Drosophila mushroom bodies
- PMID: 34107302
- PMCID: PMC8355077
- DOI: 10.1016/j.cub.2021.05.018
Sleep deprivation results in diverse patterns of synaptic scaling across the Drosophila mushroom bodies
Abstract
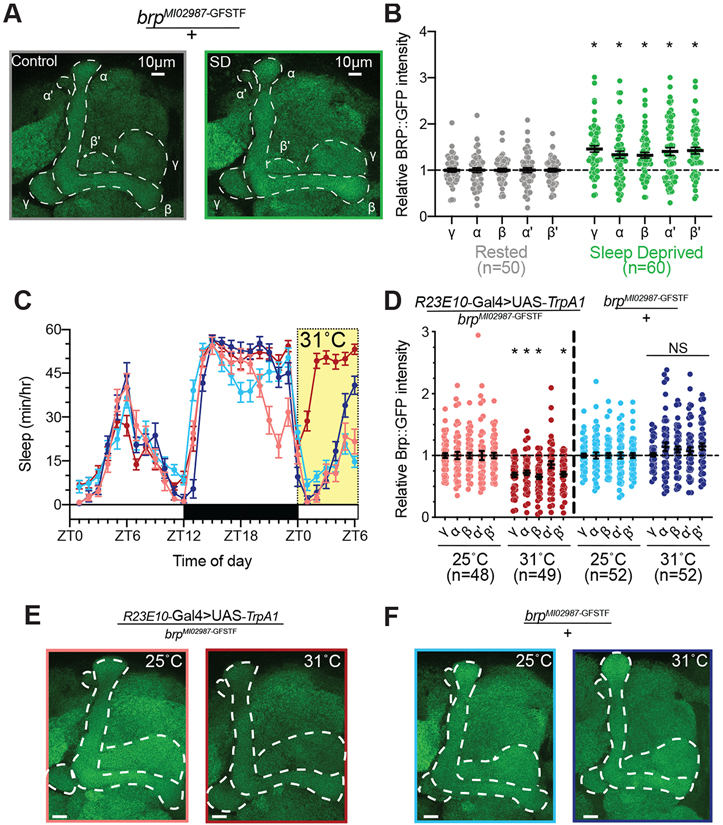
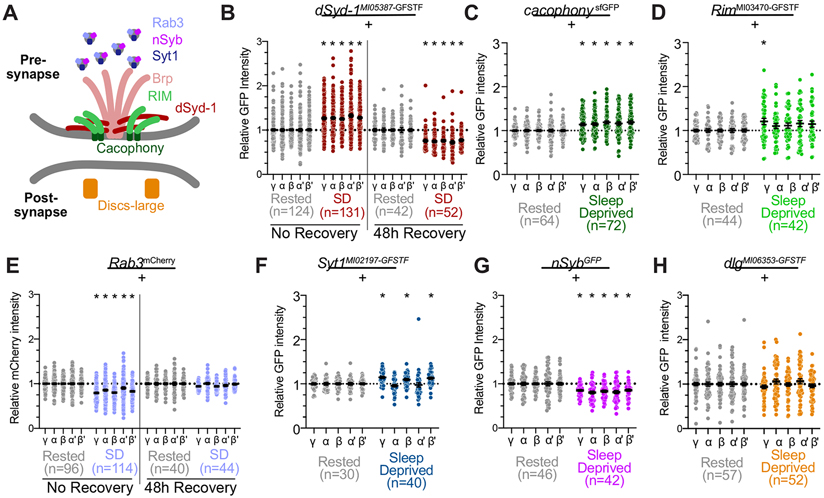
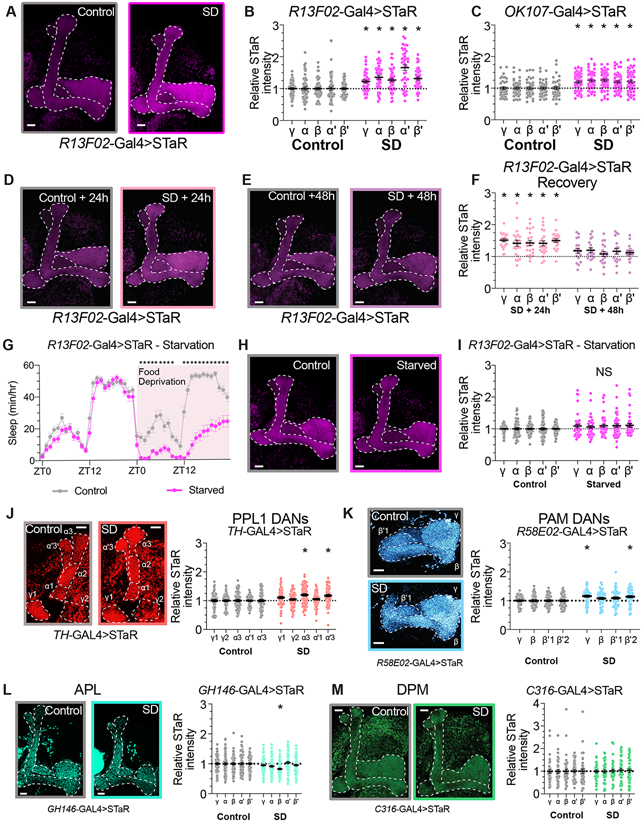
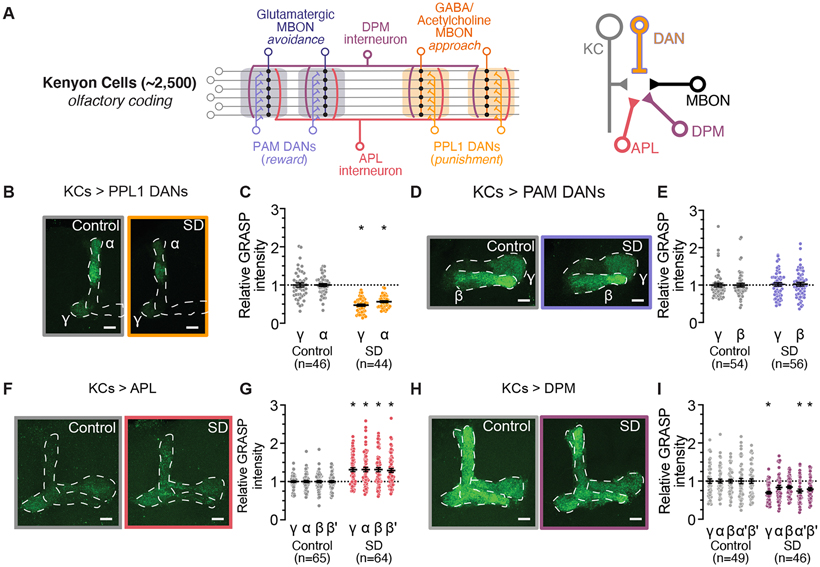
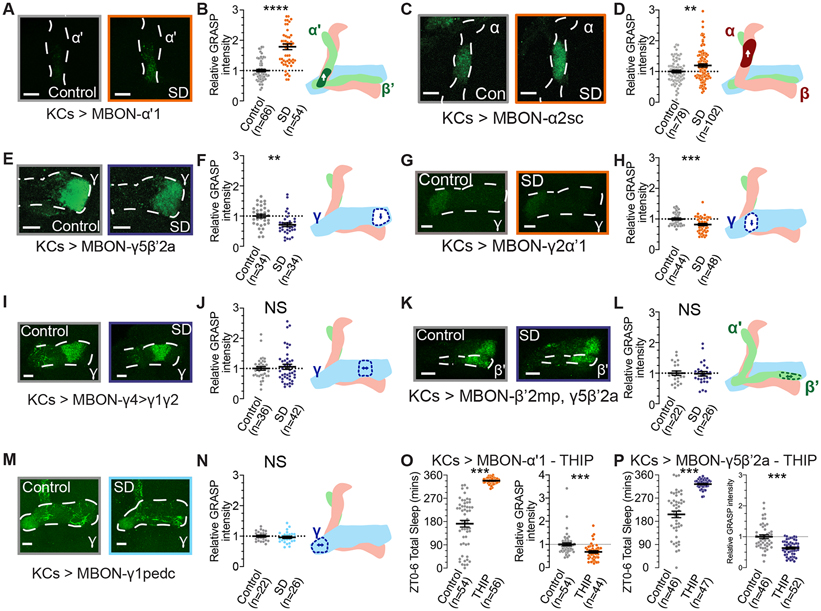
Sleep is essential for a variety of plastic processes, including learning and memory. However, the consequences of insufficient sleep on circuit connectivity remain poorly understood. To better appreciate the effects of sleep loss on synaptic connectivity across a memory-encoding circuit, we examined changes in the distribution of synaptic markers in the Drosophila mushroom body (MB). Protein-trap tags for active zone components indicate that recent sleep time is inversely correlated with Bruchpilot (BRP) abundance in the MB lobes; sleep loss elevates BRP while sleep induction reduces BRP across the MB. Overnight sleep deprivation also elevated levels of dSyd-1 and Cacophony, but not other pre-synaptic proteins. Cell-type-specific genetic reporters show that MB-intrinsic Kenyon cells (KCs) exhibit increased pre-synaptic BRP throughout the axonal lobes after sleep deprivation; similar increases were not detected in projections from large interneurons or dopaminergic neurons that innervate the MB. These results indicate that pre-synaptic plasticity in KCs is responsible for elevated levels of BRP in the MB lobes of sleep-deprived flies. Because KCs provide synaptic inputs to several classes of post-synaptic partners, we next used a fluorescent reporter for synaptic contacts to test whether each class of KC output connections is scaled uniformly by sleep loss. The KC output synapses that we observed here can be divided into three classes: KCs to MB interneurons; KCs to dopaminergic neurons; and KCs to MB output neurons. No single class showed uniform scaling across each constituent member, indicating that different rules may govern plasticity during sleep loss across cell types.
Keywords: Drosophila; active zone; bruchpilot; mushroom body; plasticity; sleep; sleep deprivation; synapse.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Walker MP, Brakefield T, Morgan A, Hobson JA, and Stickgold R (2002). Practice with sleep makes perfect: sleep-dependent motor skill learning. Neuron 35, 205–211. - PubMed
-
- Ganguly-Fitzgerald I, Donlea JM, and Shaw PJ (2006). Waking experience affects sleep need in Drosophila. Science 313, 1775–1781. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
