

Testicular germ cell-specific lncRNA, Teshl, is required for complete expression of Y chromosome genes and a normal offspring sex ratio
- PMID: 34108217
- PMCID: PMC8189594
- DOI: 10.1126/sciadv.abg5177
Testicular germ cell-specific lncRNA, Teshl, is required for complete expression of Y chromosome genes and a normal offspring sex ratio
Abstract
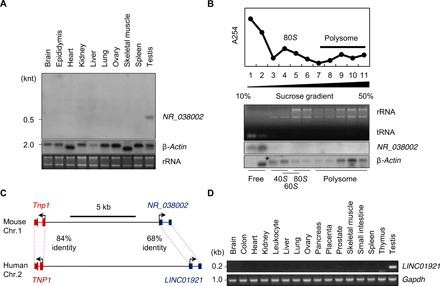
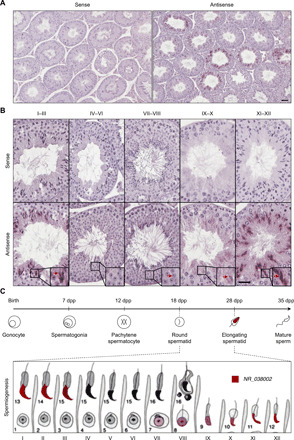
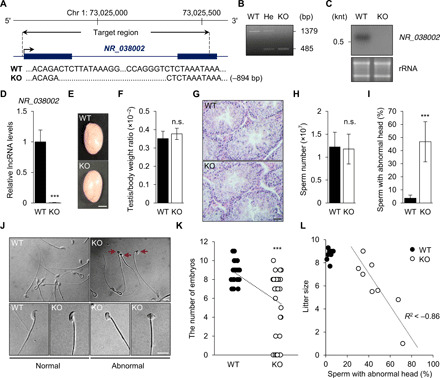
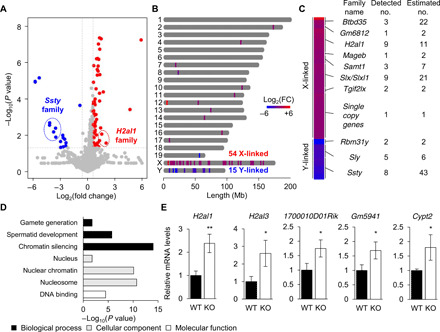
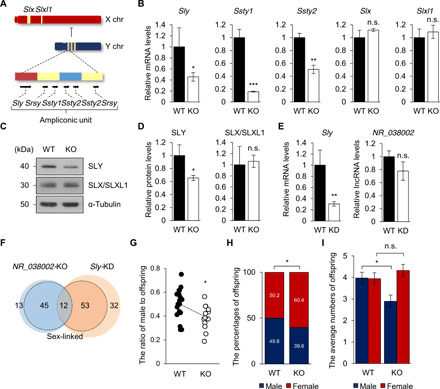
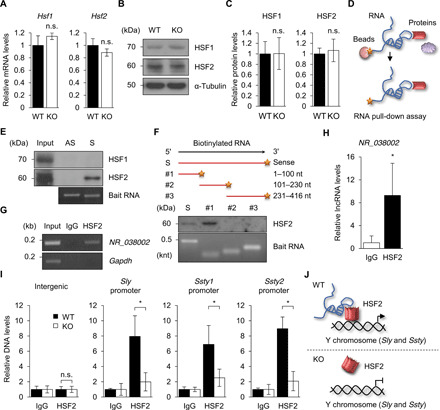
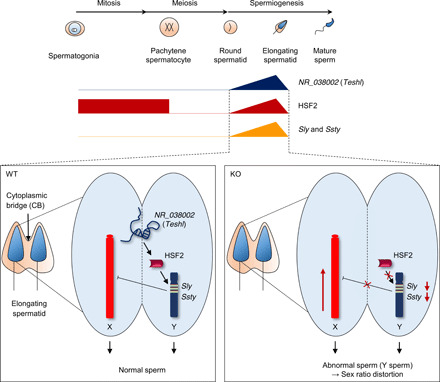
Heat shock factor 2 (HSF2) regulates the transcription of the male-specific region of the mouse Y chromosome long arm (MSYq) multicopy genes only in testes, but the molecular mechanism underlying this tissue specificity remains largely unknown. Here, we report that the testicular germ cell-specific long noncoding RNA (lncRNA), NR_038002, displays a characteristic spatiotemporal expression pattern in the nuclei of round and elongating spermatids. NR_038002-knockout male mice produced sperm with abnormal head morphology and exhibited reduced fertility accompanied by a female-biased sex ratio in offspring. Molecular analyses revealed that NR_038002 interacts with HSF2 and thereby activates expression of the MSYq genes. We designate NR_038002 as testicular germ cell-specific HSF2-interacting lncRNA (Teshl). Together, our study is the first to demonstrate that the testis specificity of HSF2 activity is regulated by the lncRNA Teshl and establishes a Teshl-HSF2-MSYq molecular axis for normal Y-bearing sperm qualities and consequent balanced offspring sex ratio.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution NonCommercial License 4.0 (CC BY-NC).
Figures
References
-
- Kapranov P., Cheng J., Dike S., Nix D. A., Duttagupta R., Willingham A. T., Stadler P. F., Hertel J., Hackermuller J., Hofacker I. L., Bell I., Cheung E., Drenkow J., Dumais E., Patel S., Helt G., Ganesh M., Ghosh S., Piccolboni A., Sementchenko V., Tammana H., Gingeras T. R., RNA maps reveal new RNA classes and a possible function for pervasive transcription. Science 316, 1484–1488 (2007). - PubMed
-
- Fatica A., Bozzoni I., Long non-coding RNAs: New players in cell differentiation and development. Nat. Rev. Genet. 15, 7–21 (2014). - PubMed
-
- Necsulea A., Soumillon M., Warnefors M., Liechti A., Daish T., Zeller U., Baker J. C., Grutzner F., Kaessmann H., The evolution of lncRNA repertoires and expression patterns in tetrapods. Nature 505, 635–640 (2014). - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
