

New substrates and determinants for tRNA recognition of RNA methyltransferase DNMT2/TRDMT1
- PMID: 34110975
- PMCID: PMC8632113
- DOI: 10.1080/15476286.2021.1930756
New substrates and determinants for tRNA recognition of RNA methyltransferase DNMT2/TRDMT1
Abstract
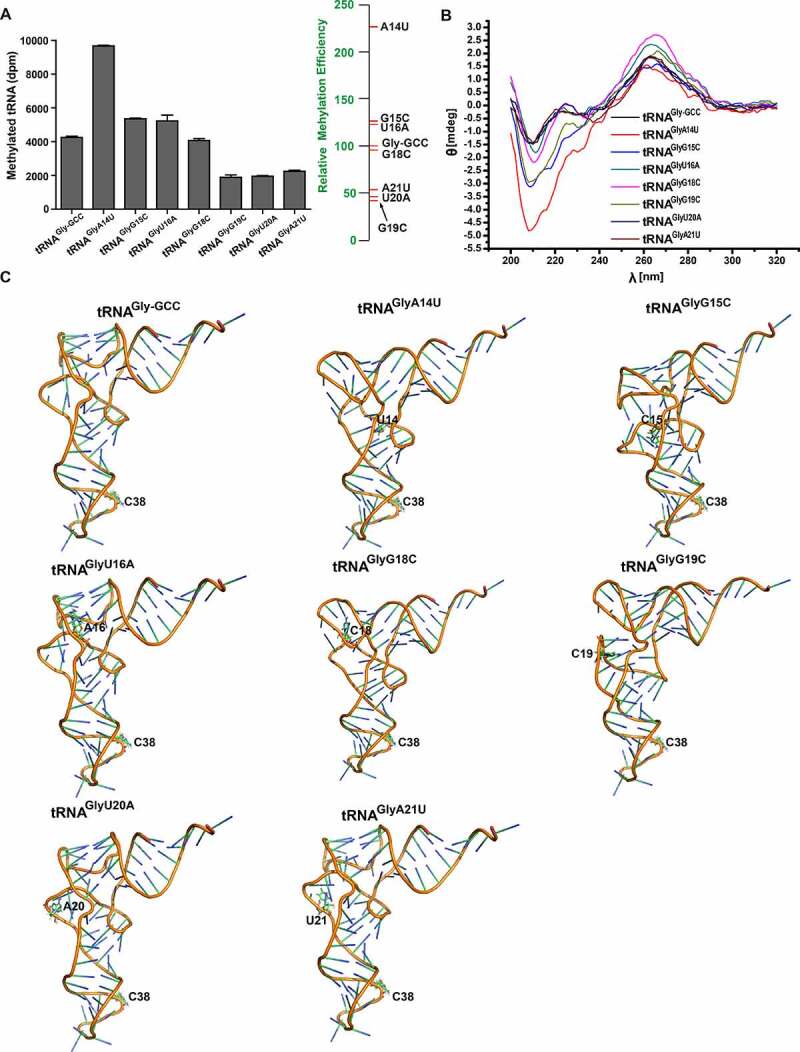
Methylation is a common post-transcriptional modification of tRNAs, particularly in the anticodon loop region. The cytosine 38 (C38) in tRNAs, such as tRNAAsp-GUC, tRNAGly-GCC, tRNAVal-AAC, and tRNAGlu-CUC, can be methylated by human DNMT2/TRDMT1 and some homologs found in bacteria, plants, and animals. However, the substrate properties and recognition mechanism of DNMT2/TRDMT1 remain to be explored. Here, taking into consideration common features of the four known substrate tRNAs, we investigated methylation activities of DNMT2/TRDMT1 on the tRNAGly-GCC truncation and point mutants, and conformational changes of mutants. The results demonstrated that human DNMT2/TRDMT1 preferred substrate tRNAGly-GCC in vitro. L-shaped conformation of classical tRNA could be favourable for DNMT2/TRDMT1 activity. The complete sequence and structure of tRNA were dispensable for DNMT2/TRDMT1 activity, whereas T-arm was indispensable to this activity. G19, U20, and A21 in D-loop were identified as the important bases for DNMT2/TRDMT1 activity, while G53, C56, A58, and C61 in T-loop were found as the critical bases. The conserved CUXXCAC sequence in the anticodon loop was confirmed to be the most critical determinant, and it could stabilize C38-flipping to promote C38 methylation. Based on these tRNA properties, new substrates, tRNAVal-CAC and tRNAGln-CUG, were discovered in vitro. Moreover, a single nucleotide substitute, U32C, could convert non-substrate tRNAAla-AGC into a substrate for DNMT2/TRDMT1. Altogether, our findings imply that DNMT2/TRDMT1 relies on a delicate network involving both the primary sequence and tertiary structure of tRNA for substrate recognition.
Keywords: DNMT2/TRDMT1; conformational change; methylation kinetics; recognition determinant; tRNA.
Figures






Similar articles
-
Determinants of DNMT2/TRDMT1 preference for substrates tRNA and DNA during the evolution.RNA Biol. 2023 Jan;20(1):875-892. doi: 10.1080/15476286.2023.2272473. Epub 2023 Nov 15. RNA Biol. 2023. PMID: 37966982 Free PMC article.
-
Position 34 of tRNA is a discriminative element for m5C38 modification by human DNMT2.Nucleic Acids Res. 2021 Dec 16;49(22):13045-13061. doi: 10.1093/nar/gkab1148. Nucleic Acids Res. 2021. PMID: 34871455 Free PMC article.
-
Restricted tRNA methylation by intermolecular disulfide bonds in DNMT2/TRDMT1.Int J Biol Macromol. 2023 Nov 1;251:126310. doi: 10.1016/j.ijbiomac.2023.126310. Epub 2023 Aug 12. Int J Biol Macromol. 2023. PMID: 37579906
-
TRDMT1-mediated RNA C-5 methylation as a novel target in anticancer therapy.Biochim Biophys Acta Rev Cancer. 2023 Nov;1878(6):188964. doi: 10.1016/j.bbcan.2023.188964. Epub 2023 Aug 23. Biochim Biophys Acta Rev Cancer. 2023. PMID: 37625528 Review.
-
Biological function molecular pathways and druggability of DNMT2/TRDMT1.Pharmacol Res. 2024 Jul;205:107222. doi: 10.1016/j.phrs.2024.107222. Epub 2024 May 21. Pharmacol Res. 2024. PMID: 38782147 Review.
Cited by
-
Advances in research on RNA methylation and cancer radiotherapy resistance.Front Oncol. 2025 Jul 31;15:1596541. doi: 10.3389/fonc.2025.1596541. eCollection 2025. Front Oncol. 2025. PMID: 40823093 Free PMC article. Review.
-
Comprehensive Analysis of Differentially Expressed Profiles of mRNA 5-Methylcytosine Modification in Metabolic Dysfunction-Associated Steatotic Liver Disease.Curr Issues Mol Biol. 2025 Apr 26;47(5):305. doi: 10.3390/cimb47050305. Curr Issues Mol Biol. 2025. PMID: 40699703 Free PMC article.
-
tRNA and tsRNA: From Heterogeneity to Multifaceted Regulators.Biomolecules. 2024 Oct 21;14(10):1340. doi: 10.3390/biom14101340. Biomolecules. 2024. PMID: 39456272 Free PMC article. Review.
-
Role of m5C methylation in digestive system tumors (Review).Mol Med Rep. 2025 Jun;31(6):142. doi: 10.3892/mmr.2025.13507. Epub 2025 Apr 4. Mol Med Rep. 2025. PMID: 40183387 Free PMC article.
-
TRDMT1 exhibited protective effects against LPS-induced inflammation in rats through TLR4-NF-κB/MAPK-TNF-α pathway.Animal Model Exp Med. 2022 Apr;5(2):172-182. doi: 10.1002/ame2.12221. Animal Model Exp Med. 2022. PMID: 35474613 Free PMC article.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials