

Runx1 shapes the chromatin landscape via a cascade of direct and indirect targets
- PMID: 34111109
- PMCID: PMC8219162
- DOI: 10.1371/journal.pgen.1009574
Runx1 shapes the chromatin landscape via a cascade of direct and indirect targets
Abstract
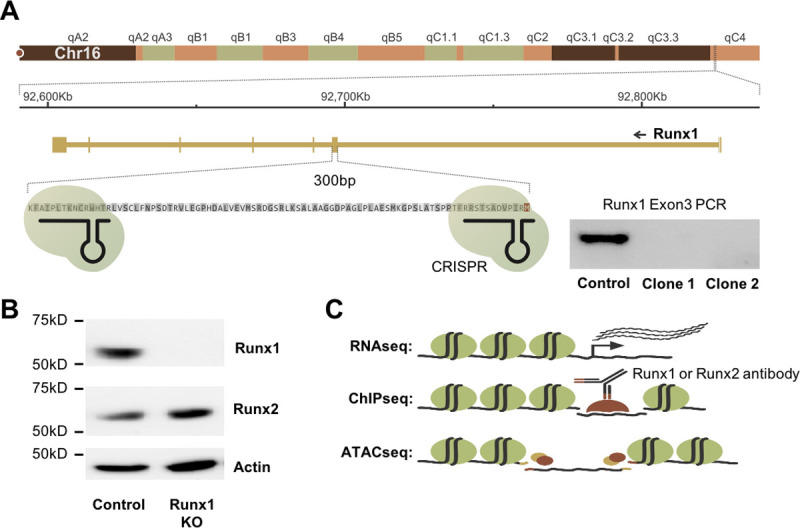
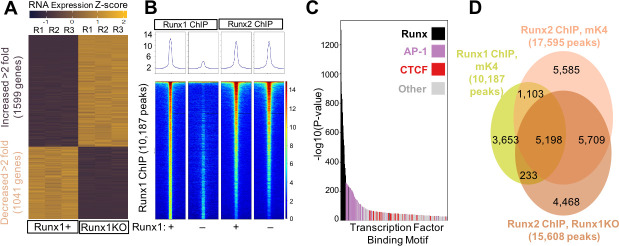
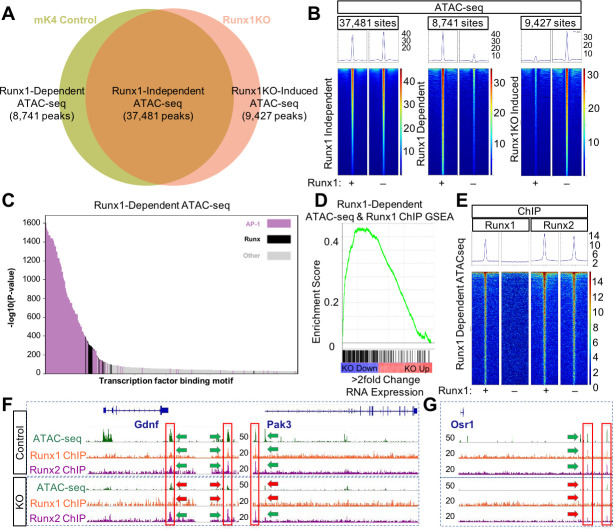
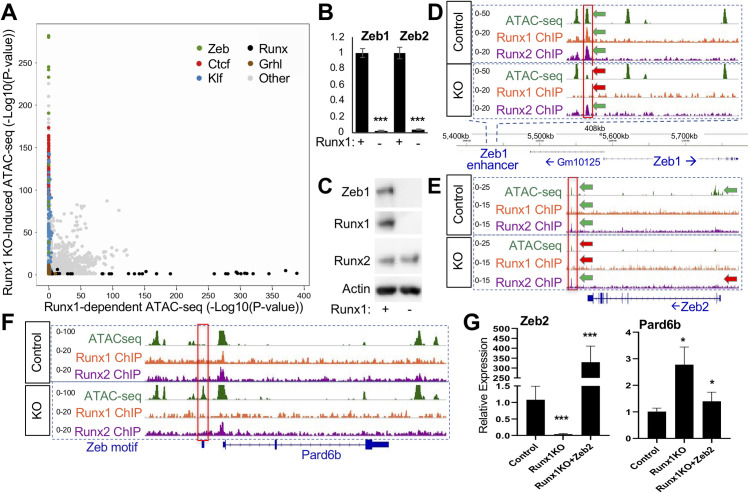
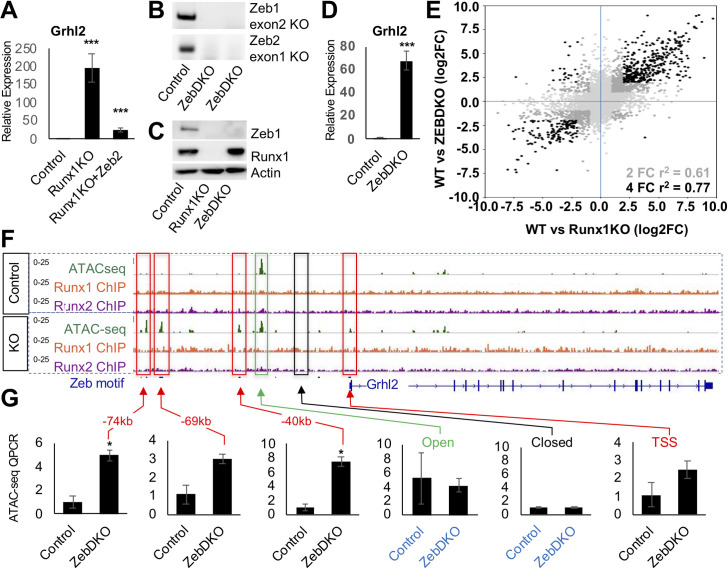
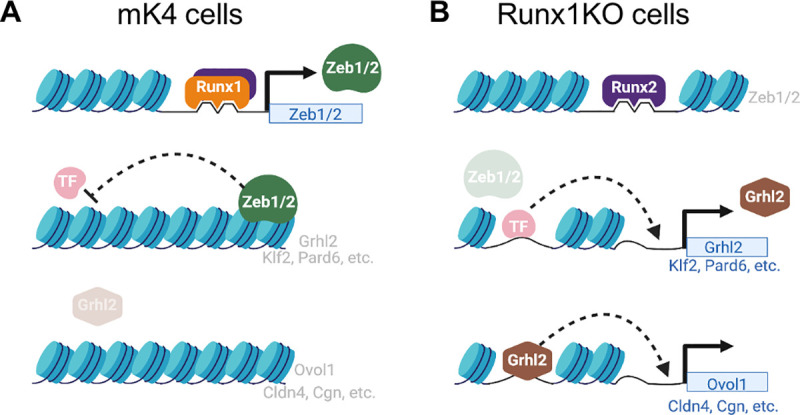
Runt-related transcription factor 1 (Runx1) can act as both an activator and a repressor. Here we show that CRISPR-mediated deletion of Runx1 in mouse metanephric mesenchyme-derived mK4 cells results in large-scale genome-wide changes to chromatin accessibility and gene expression. Open chromatin regions near down-regulated loci enriched for Runx sites in mK4 cells lose chromatin accessibility in Runx1 knockout cells, despite remaining Runx2-bound. Unexpectedly, regions near upregulated genes are depleted of Runx sites and are instead enriched for Zeb transcription factor binding sites. Re-expressing Zeb2 in Runx1 knockout cells restores suppression, and CRISPR mediated deletion of Zeb1 and Zeb2 phenocopies the gained expression and chromatin accessibility changes seen in Runx1KO due in part to subsequent activation of factors like Grhl2. These data confirm that Runx1 activity is uniquely needed to maintain open chromatin at many loci, and demonstrate that Zeb proteins are required and sufficient to maintain Runx1-dependent genome-scale repression.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Runx1 and Runx3 drive progenitor to T-lineage transcriptome conversion in mouse T cell commitment via dynamic genomic site switching.Proc Natl Acad Sci U S A. 2021 Jan 26;118(4):e2019655118. doi: 10.1073/pnas.2019655118. Proc Natl Acad Sci U S A. 2021. PMID: 33479171 Free PMC article.
-
Chromatin regulation by RUNX1.Blood Cells Mol Dis. 2010 Apr 15;44(4):287-90. doi: 10.1016/j.bcmd.2010.02.009. Epub 2010 Mar 2. Blood Cells Mol Dis. 2010. PMID: 20194037
-
RUNX1 (AML-1) and RUNX2 (AML-3) cooperate with prostate-derived Ets factor to activate transcription from the PSA upstream regulatory region.J Cell Biochem. 2006 Jan 1;97(1):1-17. doi: 10.1002/jcb.20664. J Cell Biochem. 2006. PMID: 16237704
-
Runx1 Structure and Function in Blood Cell Development.Adv Exp Med Biol. 2017;962:65-81. doi: 10.1007/978-981-10-3233-2_5. Adv Exp Med Biol. 2017. PMID: 28299651 Review.
-
RUNX1 translocations in malignant hemopathies.Anticancer Res. 2009 Apr;29(4):1031-7. Anticancer Res. 2009. PMID: 19414342 Review.
Cited by
-
Epigenetic and transcriptional dysregulation in CD4+ T cells in patients with atopic dermatitis.PLoS Genet. 2022 May 16;18(5):e1009973. doi: 10.1371/journal.pgen.1009973. eCollection 2022 May. PLoS Genet. 2022. PMID: 35576187 Free PMC article.
-
RUNX regulated immune-associated genes predicts prognosis in breast cancer.Front Genet. 2022 Aug 26;13:960489. doi: 10.3389/fgene.2022.960489. eCollection 2022. Front Genet. 2022. PMID: 36092942 Free PMC article.
-
Emerging RUNX2-Mediated Gene Regulatory Mechanisms Consisting of Multi-Layered Regulatory Networks in Skeletal Development.Int J Mol Sci. 2023 Feb 3;24(3):2979. doi: 10.3390/ijms24032979. Int J Mol Sci. 2023. PMID: 36769300 Free PMC article. Review.
-
In the spotlight: the role of TGFβ signalling in haematopoietic stem and progenitor cell emergence.Biochem Soc Trans. 2022 Apr 29;50(2):703-712. doi: 10.1042/BST20210363. Biochem Soc Trans. 2022. PMID: 35285494 Free PMC article. Review.
-
Runt-related Transcription Factors and Gene Regulatory Mechanisms in Skeletal Development and Diseases.Curr Osteoporos Rep. 2023 Oct;21(5):485-492. doi: 10.1007/s11914-023-00808-4. Epub 2023 Jul 12. Curr Osteoporos Rep. 2023. PMID: 37436583 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
